Biodiversitätserhebungen in Kräuteranbauflächen
Biodiversity surveys in medicinal and aromatic plant fields
Rilevamenti della biodiversità in campi di erbe medicinali e aromatiche
1 EURAC Research, 39100 Bozen/Bolzano, Italy
2 Laimburg Research Centre, 39040 Auer/Ora, BZ, Italy
3 no affiliation
4 Fondazione Edmund Mach, Centro Trasferimento Tecnologico, 38098 San Michele all’Adige, TN, Italy
5 Università degli Studi di Milano, 20133 Milano, Italy
6 Universität Innsbruck, 6020 Innsbruck, Austria
Abstract
Medicinal and aromatic plants (MAPs) in mountain regions are cultivated on small-scale farms, which are characterized by a great diversity of MAPs grown on a relatively small area and by a high degree of habitat complexity. Non-crop elements (e.g., dry-stone walls, hedges, etc.) are widely present in these cultivated areas and, together with high plant diversity, may provide ideal foraging and breeding habitats for several animal groups. Here we surveyed small-scale MAP fields from a multi-taxonomic perspective considering flower-visiting arthropods, butterflies, grasshoppers, ground-dwelling arthropods, birds, and bats. A total of three MAP fields were surveyed, however not every taxon was surveyed in each MAP field. Pan traps were used in all MAP fields to assess flower-visiting arthropods with special attention to wild bees. In one of the selected fields a Malaise trap was used, and the other taxa were surveyed according to the protocol of the Biodiversity Monitoring South Tyrol. An exception was the bird surveys, which were conducted in two MAP fields. Our results indicate MAP fields to be a valuable habitat for several taxa, especially wild bees, as reflected in the positive correlation of wild bee species richness and flower coverage. Next to beneficial arthropods, potential pests such as aphids were also highly abundant. However, natural enemies (e.g., hymenopteran parasitoids, ground-dwelling predators, etc.) were also numerous and possibly counteracted pests. The butterfly and grasshopper fauna were represented by common and generalist species, while the observed vertebrate communities were relatively diverse in their habitat requirements, most likely using MAP fields for foraging. Overall, we conclude that MAP cultivation sites may act as resource-rich oases for several animal groups, thereby also promoting biodiversity on a broader scale.
Arznei- und Gewürzpflanzen werden in Bergregionen in kleinen Kräuteranbau-Betrieben, die sich durch eine hohe Pflanzen- und Strukturvielfalt (Trockenmauern, Hecken, usw.) auszeichnen, angebaut. In dieser Studie wurden Kräuteranbau-Betriebe als Lebensraum für verschiedene Tiergruppen untersucht, wobei blütenbesuchende Arthropoden (Wildbienen, Bock- und Prachtkäfer), Schmetterlinge, Heuschrecken, bodenoberflächen-aktive Arthropoden, Vögel und Fledermäuse berücksichtigt wurden. Die Kräuteranbau-Betriebe stellten sich als ein wertvoller Lebensraum für verschiedene Tiergruppen heraus, die möglicherweise die Flächen als Nahrungs- oder auch als Bruthabitat nutzen. Insbesondere Wildbienen waren mit 10% des regionalen Artenpools besonders zahlreich und reagierten positiv auf das Vorkommen von Blüten. Neben nützlichen Arthropoden waren auch potenzielle Schädlinge sehr häufig anzutreffen, wobei natürliche Feinde, wie Räuber und Parasitoide, ebenfalls zahlreich vertreten waren. Die Schmetterlings- und Heuschreckenfauna war durch häufige und generalistische Arten vertreten, während Vögel und Fledermäuse sich durch verschiedene Lebensraum- und Landschaftsansprüche auszeichneten. Insgesamt können Kräuteranbau-Betriebe in der Agrarlandschaft als ressourcen- und strukturreiche Oasen für verschiedene Tiergruppen fungieren und sich somit womöglich auch positiv auf die Biodiversität auf einer breiteren Skala auswirken.
Le erbe medicinali ed aromatiche nelle regioni montane vengono coltivate in piccole aziende agricole. Queste sono caratterizzate da un'elevata diversità di piante coltivate e da un elevato grado di complessità della struttura dell’habitat dovuta alla presenza di elementi naturali e semi-naturali non colturali (es. muri a secco, siepi, ecc.). In questo studio sono stati analizzati alcuni gruppi di animali presenti in tre campi di erbe medicinali e aromatiche differenti. Nello specifico sono stati rilevati insetti visitatori di fiori (api selvatiche, cerambicidi e buprestidi), farfalle diurne, cavallette, artropodi della superficie del suolo, uccelli e pipistrelli. I risultati indicano che i campi di erbe medicinali ed aromatiche sono un habitat prezioso per diversi taxa, i quali usano questo ambiente per l'alimentazione o per la riproduzione. I campi di erbe sono soprattutto importanti per le api selvatiche, come dimostra la correlazione positiva tra la ricchezza di specie di api selvatiche e la percentuale di copertura floreale. Inoltre, sono state rilevate circa il 10% delle specie di api selvatiche presenti nella provincia, ciò evidenzia che questi habitat sono particolarmente ricchi di specie. Oltre agli insetti impollinatori, anche insetti potenzialmente dannosi per l’agricoltura, come gli afidi, e insetti antagonisti naturali come predatori e parassitoidi sono risultati abbondanti. Le comunità di farfalle e cavallette sono caratterizzate da specie comuni e generaliste, mentre per gli uccelli e pipistrelli sono state rilevate specie esigenti per qualità di habitat e paesaggio che probabilmente utilizzano i campi di erbe per il foraggiamento. Nel complesso, i campi di erbe medicinali ed aromatiche possono fungere nell’ambiente agricolo da oasi ricche di risorse e strutture naturali per diversi gruppi di animali, promuovendo un alto valore di biodiversità anche ad una scala più ampia.
Introduction
Medicinal and aromatic plants (MAPs) are an important part of our natural wealth and have long been used to cure diseases and increase human wellbeing [1]. About two-thirds of the world population in developing countries rely on plant-based traditional medicines for primary healthcare requirements [2]. The popularity and acceptance of MAPs can also be observed in industrialized countries, where derived products find use in homeopathy, phytotherapy and aromatherapy [1]. In the United States the market for herbal supplements alone is worth nearly $4 billion per year [1]. MAPs have gained importance in industrial production as raw materials for the manufacture of various products (e.g., essential oils, dyes, biocides, cosmetics, etc.) and in the food industry in the form of spices, condiments, food preservatives and additives [1][3]. For these purposes, an estimated number of 50 000 plant species worldwide are used as MAPs, of which approximately two thirds are wild harvested, and the rest are cultivated [1][4][5]. In Europe, MAPs are grown on an area of more than 100,000 hectares, with France, Germany, Austria, and Finland being the largest producers with the largest cultivated areas [3]. MAPs are often grown in monocultures to ensure large quantities and high quality for industrial purposes and export markets [1][6]. Most source countries export raw crop material, while processing (value-added) takes place in consumer countries and trading centers [7]. MAP monocultures contrast with the small-scale cultivation of MAPs, where plots are usually small and treated like market gardens and small-scale arable fields [6]. This is accompanied by partly manual harvesting, on-farm processing, and marketing, which requires a high labor input and know-how that needs to be managed by small-scale farmers [6].
×
- [1] Máthé Á. (2015). Utilization/Significance of Medicinal and Aromatic Plants. In: Máthé Á (ed.). Medicinal and Aromatic Plants of the World. Scientific, Production, Commercial and Utilization Aspects. Springer, Heidelberg, Germany, here pp. 1-12, DOI: 10.1007/978-94-017-9810-5.
- [2] Ahad B., Shahri W., Rasool H. et al. (2021). Medicinal Plants and Herbal Drugs. An Overview. In: Aftab T., Hakeem K.R. (eds.). Medicinal and Aromatic Plants. Healthcare and Industrial Applications. Springer, Cham, Switzerland, here p. 2, DOI: 10.1007/978-3-030-58975-2_1.
- [3] Lubbe A., Verpoorte R. (2011). Cultivation of medicinal and aromatic plants for specialty industrial materials. Industrial Crops and Products 34 (1), 785-801, DOI: 10.1016/j.indcrop.2011.01.019.
- [4] Leaman D.J. (2008). Conservation and Sustainable Use of Wild-sourced Botanicals. Planta Medica, 74, 11, DOI: 10.1055/s-2008-1075152.
- [5] Shippmann U., Leaman D.J., Cunningham A.B. (2002). Impact of Cultivation and Gathering of Medicinal Plants on Biodiversity. Global Trends and Issues. In: Biodiversity and the Ecosystem Approach in Agriculture, Forestry and Fisheries. Satellite event on the occasion of the Ninth Regular Session of the Commission on Genetic Resources for Food and Agriculture, Rome, Italy, October 12-13, 2002. Food and Agriculture Organization of the United Nations, Rome, Italy.
- [6] Salomon I., Haban M., Otepka P. et al. (2018). Perspectives of small- and large- scale cultivation of medicinal, aromatic and spice plants in Slovakia. Medicinal Plants 10 (4), 261-267, DOI: 10.5958/0975-6892.2018.00041.2.
- [7] Allen D., Bilz M., Leaman D.J. et al. (2014). European Red List of Medicinal Plants. Publications Office of the European Union, Luxembourg, Luxembourg, here p. 4.
In South Tyrol, a mountain region in the Central Alps, MAP cultivation represents a small but culturally important part of South Tyrolean agriculture, with a total of 47 farmers cultivating an area of approximately 20 hectares. These small-scale farms, with a cultivated area of only a few 1000 m2 per farm, are mainly located in mountain areas ranging from 530 and 1700 m a.s.l. The local cultivation of MAPs encompasses a diverse array of approximately 120 annual and perennial species. Among these, the most commonly grown species include lemon balm (Melissa officinalis L.), peppermint (Mentha x piperita), sage (Salvia officinalis L.), thyme (Thymus vulgaris L.), oregano (Origanum vulgare L.), marjoram (Origanum majorana L.), and common nettle (Urtica dioica L.). In general, most of these species are harvested either before or during the onset of the flowering period to ensure optimal MAP properties. Flowers such as marigolds (Calendula officinalis L.), cornflowers (Centaurea cyanus L.), and lavender (Lavandula angustifolia Mill.) are also grown ([8], Manuel Pramsohler pers. comm.).
×
- [8] Südtiroler Bauernbund (ed.) (2014). Nischenkulturen als Erwerbsmöglichkeit. Chance und Herausforderung für die Südtiroler Landwirtschaft. Südtiroler Bauernbund, Bozen/Bolzano, Italy. Retrieved December 15, 2022, from https://issuu.com/effektgmbh/docs/sbb_broschu__re_nischenkulturen.
The small-scale cultivation of MAPs in a mountain region like South Tyrol suggests that MAP fields are of high ecological value [9]. Fields are rich in non-crop structures, such as hedges, fences, and dry-stone walls, and may provide continuous and diverse food resources for arthropods and other animals. Also, Licata et al. (2022) [10] stated that sustainable and integrated MAP cultivation can aid in maintaining and enhancing the biodiversity of agroecosystems. However, research on this topic is generally lacking. Most zoology related studies that have been conducted in MAP fields have focused on harmful arthropods [11], such as beetles (i.e., leaf beetles, weevils, etc.) [12], leafhoppers [13][14], aphids, phytophagous thrips [14][15] and mites [16]. Only a few studies have addressed other insect groups, such as pollinators and their contribution to the species-specific reproductive success of MAPs [17][18]. Studies that focus on other invertebrate and vertebrate groups in MAP fields and that attempt to determine biodiversity from a multi-taxonomic perspective are lacking. To close this gap, we surveyed several taxa in MAP fields, mainly focusing on arthropods (with a special focus on flower-visiting insects, incl. pollinator and pest species), but also on the avian and bat fauna.
×
- [9] Šálek M., Hula V., Kipson M. et al. (2018). Bringing diversity back to agriculture. Smaller fields and non-crop elements enhance biodiversity in intensively managed arable farmlands. Ecological Indicators 90, 65-73, DOI: 10.1016/j.ecolind.2018.03.001.
- [10] Licata M., Maggio A.M., La Bella S. et al. (2022). Medicinal and aromatic plants in agricultural research, when considering criteria of multifunctionality and sustainability. Agriculture 12 (4), 529, DOI: 10.3390/agriculture12040529.
- [11] Hoppe B. (ed.) (2007). Handbuch des Arznei- und Gewürzpflanzenanbaus. Bd. 3: Krankheiten und Schädigungen an Arznei- und Gewürzpflanzen. SALUPLANTA e. V., Bernburg, Germany.
- [12] Pramsohler M., Gallmetzer A., Castellan A. et al. (2022). Erster Nachweis und molekularbiologische Bestimmung von Donus intermedius (Coleoptera: Curculionoidae) als Schädling bei Zitronenmelisse in Südtirol. Laimburg Journal 4, DOI: 10.23796/LJ/2022.003.
- [13] Nickel H., Blum H., Jung K. (2014). Verbreitung und Biologie der an mitteleuropäischen Arznei- und Gewürzpflanzen schädlichen Blattzikaden (Hemiptera: Cicadellidae, Typhlocybinae). Cicadina 14, 13-42, DOI: 10.25673/92235.
- [14] Meyer U., Blum H., Gärber U. et al. (2010). Praxisleitfaden Krankheiten und Schädlinge im Arznei- und Gewürzpflanzenanbau. Julius-Kühn-Institut Selbstverlag, Braunschweig, Germany.
- [15] Pobożniak M., Sobolewska A. (2011). Biodiversity of thrips species (Thysanoptera) on flowering herbs in Cracow, Poland. Journal of Plant Protection Research 51 (4), 393-398, DOI: 10.2478/v10045-011-0064-2.
- [16] Gupta S.K., Karmakar K. (2011). Diversity of mites (Acari) on medicinal and aromatic plants in India. Zoosymposia 6, 56-61, DOI: 10.11646/zoosymposia.6.1.10.
- [17] Venkatesh Y.N., Neethu T., Ashajyothi M. et al. (2022). Pollinator activity and their role on seed set of medicinal and aromatic Lamiaceae plants. Journal of Apicultural Research, Vol. ahead-of-print, DOI: 10.1080/00218839.2022.2080949.
- [18] Kumari B., Ravinder R. (2017). Pollination studies in Tagetes minuta, an important medicinal and aromatic plant. Medicinal Plants – International Journal of Phytomedicines and Related Industries 9 (2), 140-142, DOI: 10.5958/0975-6892.2017.00021.1.
Methods
Research area and study sites
The study was conducted from May to August 2021 in the Autonomous Province of South Tyrol, Italy. The study sites were located on three small-scale farms specialized in organic cultivation of medicinal and aromatic plants (MAP) (Fig. 1a). The study sites were located in Merano/Meran (M) (620 m a.s.l.; 46°39’59.213”N, 11°11’45.672”E), in Castelrotto/Kastelruth (K) (800 m a.s.l., 46°32’47.698”N, 11°31’35.288”E) and in Prati/Wiesen (W) (980 m a.s.l., 46°54’9.468”N, 11°28’11.765”E). Data was collected in the framework of the Biodiversity Monitoring South Tyrol project [19] in collaboration with Laimburg Research Centre.
×
- [19] Hilpold A., Anderle M., Guariento E. et al. (2023). Handbook - Biodiversity Monitoring South Tyrol. Eurac Research, Bolzano, Italy, DOI: 10.57749/2qm9-fq40.

Data collection
Pan traps
Yellow, white, and blue pan traps (PT) were used to collect flower-visiting arthropods in all three MAP fields during three survey events (Fig. 1b). Survey events occurred once per month from mid-June to late August. PT were made using a round saucer 16 cm in diameter and with a height of 2 cm (01 011 016, TeraPlast S.p.A., Castelgomberto, Italy). PT were dyed with UV-reflecting yellow paint, blue paint, and white paint after white priming. PT were deployed in sets of three. Each set consisted of one PT of each color. During each survey event four sets of PT were deployed in M and K, and five sets in W, resulting in 12 PT in M and K and 15 PT in W per survey event. PT were placed specifically in areas of high crop diversity and their heights were adjusted to the level of the surrounding vegetation [20]. The additional fifth PT set in W (PT B) represented an exception, as it was surrounded by only one plant species. The cover of each MAP species was recorded in a quadrat of 5 x 5 m around every PT set. The phenology (NF = no flower, BF = beginning of flowering period, F = flower, EF = end of flowering period) of each MAP species was assessed per survey event. A distance of at least 5 m was maintained between each set of PT [21]. PT were filled with water and a drop of unscented dish soap. After 24 hours, collected arthropods were transferred to 70% Ethanol.
×
- [20] Vrdoljak S.M., Samways M.J. (2010). Optimising coloured pan traps to survey flower visiting insects. Journal of Insect Conservation 16, 345-354, DOI: 10.1007/s10841-011-9420-9.
- [21] Droege S., Tepedino V.J., Lebuhn G. et al. (2010). Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conservation and Diversity 3 (1), 15-23, DOI: 10.1111/j.1752-4598.2009.00074.x.

The sorting of arthropods was performed by the authors LO and LN. Arthropods were sorted according to order level and for Collembola by class level. For some arthropod groups (Diptera, Coleoptera, Hymenoptera, Hemiptera and Arachnidae) further identification to lower taxonomic levels was performed. Within Coleoptera, Cerambycidae and Buprestidae, specimens were identified to the species level by GvM. Parasitoids of the Hymenoptera order were identified down to the family level by LO using the key in Goulet and Huber (1993) [22]. Exceptions were made for Cynipoidea, which were identified to the superfamily level. Some representatives of the Cynipoidea are gall-forming wasps [22], which is why this superfamily was not counted as parasitoids. For some families of Chalcidoidea, identification down to the family level was not possible. Therefore, the respective families were summarized as "Chalcidoidea varia". Furthermore, within the Braconidae family the Aphidiinae subfamily was separated. Wild bees, which include all bee species of the Apoidea superfamily with exception of the domesticated Apis mellifera (Linnaeus, 1758), were identified to the species level by PF and team.
×
- [22] Goulet H., Huber J.T. (eds.) (1993). Hymenoptera of the world. An identification guide to families. Canada Communication Group, Ottawa, Canada, here pp. 65-529.
Additional surveys
In M, Malaise traps were installed three times for five consecutive days from mid-June to late August. Collecting bottles were filled with 70% ethanol and transferred to fresh ethanol until further processing. Sorting and identification of arthropods was performed by LO following the same procedure as the PT sorting and identification.
Birds, bats, grasshoppers, butterflies, and ground-dwelling macro-invertebrates were surveyed following the protocol of the Biodiversity Monitoring South Tyrol [19]. Birds were surveyed in M and W, while other taxa were surveyed only in M.
×
- [19] Hilpold A., Anderle M., Guariento E. et al. (2023). Handbook - Biodiversity Monitoring South Tyrol. Eurac Research, Bolzano, Italy, DOI: 10.57749/2qm9-fq40.
Birds were recorded by MA during two survey events per MAP field using vocal point counts and visual identification [23][24]. Surveys lasted 10 minutes and considered only birds within a radius of 100 m from the selected point [25].
×
- [23] Bibby C.J., Burgess N.D., Hillis D.M. et al. (2000). Bird census techniques. Academic Press, London, United Kingdom, here pp. 91-112.
- [24] Anderle M., Paniccia C., Brambilla M. et al. (2022). The contribution of landscape features, climate and topography in shaping taxonomical and functional diversity of avian communities in a heterogeneous Alpine region. Oecologia, 199, 499-512, DOI: 10.1007/s00442-022-05134-7.
- [25] Fornasari L., Mingozzi T. (1999). Monitoraggio dell'avifauna nidificante in Italia. Un progetto pluriennale sulle specie comuni. Avocetta - Journal of Ornithology, 23, 153-153.
For surveying bat activity, an ultrasonic recording device was installed at 1.5 m above ground to record ultrasonic echolocation calls for three consecutive nights from 20 to 23 September 2021. The device was active from sunset to sunrise. Echolocation calls were identified at the species level when possible and assigned as feeding buzzes or social calls by CP [26][27].
×
- [26] Barataud M. (2015). Acoustic ecology of European bats. Species Identification and Studies of Their Habitats and Foraging Behaviour. Biotope & National Museum of Natural History, Paris, France, here pp. 104-263.
- [27] Teets K.D., Loeb S.C., Jachowski D.S. (2019). Detection probability of bats using active versus passive monitoring. Acta Chiropterologica 21 (1), 205-213, DOI: 10.3161/15081109ACC2019.21.1.017.
Grasshopper surveys were performed by AH on 28 July with a sweep net in a 10 x 10 m plot, starting with a 15 m transect along the diagonal of the plot. Additional sweep net beats, detailed hand searches and acoustic assessments were performed in the whole plot within a total of 30 minutes [28].
×
- [28] Hilpold A., Kirschner P., Dengler J. (2020). Proposal of a standardized EDGG surveying methodology for orthopteroid insects. Palaearctic Grasslands 46, 52-57, DOI: 10.21570/EDGG.PG.46.52-57.
Butterflies were surveyed five times from May to August by EG. During each survey event an area of 1000 m2 (20 x 50 m) around the central point was surveyed for 30 minutes [29].
×
- [29] Guariento E., Rüdisser J., Fiedler K. et al. (2022). From diverse to simple: butterfly communities erode from extensive grasslands to intensively used farmland and urban areas. Biodiversity and Conservation 32, 867-882, DOI: 10.1007/s10531-022-02498-3.
Ground-dwelling macro-invertebrates were collected with pitfall traps by JP. A glass jar with a diameter of 7.5 cm and a height of 9 cm was filled with 200 ml 75% propylene glycol. The traps were protected from rain and other disturbances by a polycarbonate Lexan® roof. Two traps per survey period were set, once in early summer and once in autumn, resulting in a total of four traps exposed for 15 days each.
Analysis
Descriptive analyses were performed without the additional fifth PT in W. However, total abundances including PT B are reported for completeness in the tables. Species accumulation curves were calculated for wild bees to represent differences in diversity and sample completeness in M, K, and W (- PT B) (R version 4.0.4, package iNEXT) [30]. Correlations between flower abundance (cover of flowering MAP/per survey event) and the abundance of Syrphidae, A. mellifera, wildbees and wildbee species richness, were tested using Pearson correlation (R version 4.0.4, package ggplot2 and ggpubr).
×
- [30] Hsieh T.C., Ma K.H., Chao A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods of Ecology and Evolution 7 (12), 1451-1456, DOI: 10.1111/2041-210X.12613.
Results
Medicinal and aromatic plants around the pan traps
A total of 38 species and two genera of medicinal and aromatic plants (MAP) from 11 different plant families were recorded around the pan traps (PT) (Fig. 2). In Merano/Meran (M) and Prati/Wiesen (W), 17 MAP species were recorded each, and 21 recorded in Castelrotto/Kastelruth (K). The MAP species, which accounted for 50% of the cover around the PT, were Mentha sp. (12.4%), Monarda didyma L. (10.7%), Lavandula angustifolia (8.6%), Thymus citriodorus (Pers.) Schreb. (8.6%), Nepeta cataria L. (6.5%), and Calendula officinalis L. (5.3%). The MAP species recorded in each MAP field are listed in Table 1.
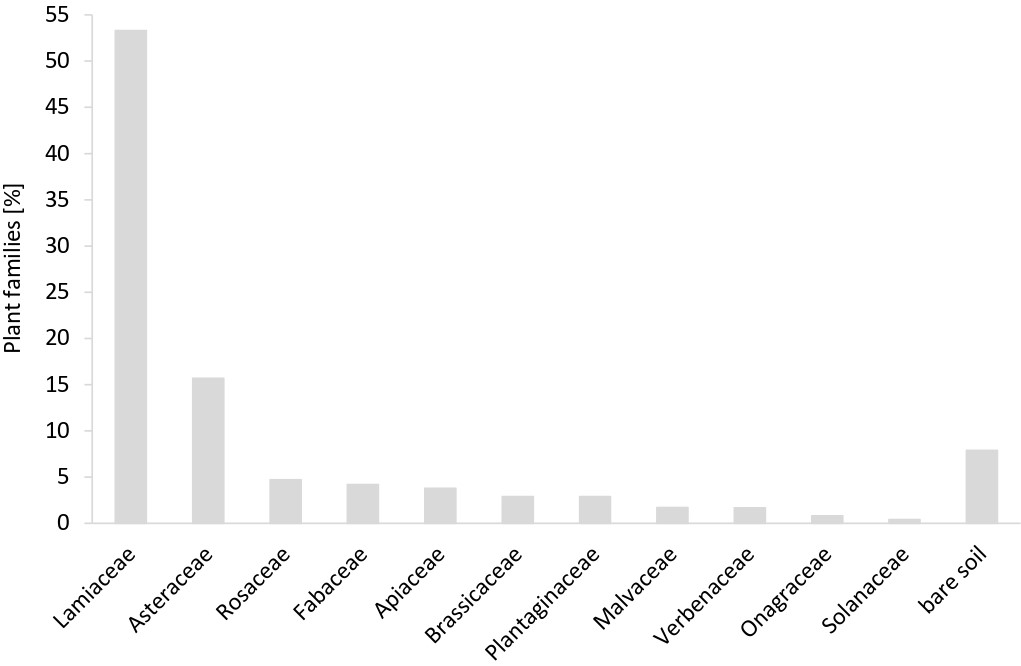
+
Fig. 2: Percentage of medicinal and aromatic plant families recorded in quadrats of 5 x 5 m around each pan trap set.
Tab. 1: Species list of the medicinal and aromatic plant (MAP) species recorded around the pan traps per MAP field (M = Merano/Meran, K = Kastelruth/Castelrotto, W = Wiesen/Prati). The number 1 represents the presence of the MAP in the respective MAP field.
+
Taxa |
M |
K |
W |
|---|---|---|---|
Achillea millefolium L. |
|
1 |
|
Agastache anethiodora (Nutt.) Britton |
|
1 |
|
Alcea rosea L. |
1 |
|
|
Alchemilla vulgaris L. |
1 |
1 |
|
Althaea officinalis L. |
1 |
|
|
Arnica montana L. |
1 |
|
|
Artemisia dracunculus L. |
|
1 |
|
Calendula officinalis L. |
1 |
1 |
1 |
Carumcarvi L. |
|
1 |
|
Centaurea cyanus L. |
1 |
1 |
1 |
Coriandrum sativum L. |
|
1 |
|
Dracocephalum moldavica L. |
|
1 |
|
Echinacea purpurea (L.) Moench |
|
1 |
|
Echinacea sp. |
1 |
|
|
Fagopyrum esculentum Moench. |
|
1 |
|
Hyssopus officinalis L. |
|
1 |
|
Lavandula angustifolia MILL. |
1 |
1 |
|
Leontopodium alpinum Cass. |
|
1 |
|
Malva sylvestris L. |
1 |
1 |
|
Matricaria chamomilla L. |
1 |
|
|
Melissa officinalis L. |
1 |
1 |
1 |
Mentha sp. |
|
1 |
1 |
Mentha spicata L. |
|
1 |
|
Monarda didyma L. |
1 |
1 |
1 |
Monarda fistulosa L. |
|
1 |
|
Nepeta cataria L. |
|
1 |
1 |
Ocimum basilicum L. |
1 |
|
|
Oenothera biennis L. |
|
1 |
|
Origanum majoran L. |
|
1 |
|
Origanum vulgare L. |
|
1 |
|
Petroselinum crispum (Mill.) Fuss |
1 |
|
|
Plantago lanceolata L. |
|
1 |
|
Sanguisorba minor Scop. |
1 |
|
|
Satureja montana L. |
|
1 |
|
Sinapis alba/Raphanus sativus |
1 |
|
|
Solanum tuberosum L. |
|
1 |
|
Thymus citriodorus (Pers.) Schreb. |
1 |
1 |
|
Trigonella caerulea L. |
|
1 |
|
Urtica dioica L. |
|
1 |
|
Verbena officinalis L. |
|
1 |
|
Veronica sp. |
|
1 |
|
Species richness |
17 |
21 |
17 |
Pan traps
Overall, 12 570 arthropods were collected with PT, of which 12 155 were considered target taxa attracted by the colors of the PT (flower visiting Coleoptera, Diptera, Hymenoptera, Thysanoptera, Lepidoptera, Sternorrhyncha). An average of 4051.7 ± 684.7 target taxa were recorded per MAP field. The total abundance and average abundance of each taxon per MAP field is listed in Table 2.
Tab. 2: Abundance of taxa recorded with Malaise traps (MT) in Merano/Meran (M) and with pan traps (PT) in each medicinal and aromatic plant field (MAP) (M, K = Castelrotto/Kastelruth, W = Prati/Wiesen). Taxa recorded with PT are divided into target taxa of PT and non-target taxa of PT. For W, values with (+ PT B) and without (- PT B) the additional pan trap set are indicated. Average abundance (± SD) per MAP field and taxon were calculated with M, K, and W (- PT B).
+
Taxa |
MT |
PT |
||||||
|---|---|---|---|---|---|---|---|---|
Abundance |
Abundance |
M |
K |
W |
W |
Average (± SD)/MAP field |
||
(- PT B) |
(+ PT B) |
|||||||
Target taxa of PT |
Coleoptera varia |
102 |
524 |
247 |
188 |
89 |
101 |
174.7 (SD ± 79.8) |
Buprestidae |
89 |
12 |
53 |
24 |
28 |
29.7 (SD ± 21.1) |
||
Cerambycidae |
2 |
29 |
4 |
5 |
20 |
25 |
9.7 (SD ± 9) |
|
Curculionidae |
14 |
7 |
5 |
1 |
1 |
1 |
2.3 (SD ± 2.3) |
|
Diptera varia |
1004 |
7585 |
1303 |
2686 |
3596 |
4259 |
2528.3 (SD ± 1154.6) |
|
Syrphidae |
21 |
222 |
28 |
155 |
39 |
54 |
74 (SD ± 70.4) |
|
Parasitoids/Cynipoidea |
514 |
334 |
123 |
138 |
73 |
90 |
111.3 (SD ±34) |
|
Wild bees |
20 |
235 |
67 |
91 |
77 |
81 |
78.3 (SD ± 12.1) |
|
Apis mellifera |
2 |
151 |
74 |
57 |
20 |
30 |
50.3 (SD ± 27.6) |
|
Symphyta |
11 |
100 |
4 |
16 |
80 |
101 |
33.3 (SD ± 40.9) |
|
Vespoidea/Spheciformes |
31 |
86 |
29 |
14 |
43 |
52 |
28.7 (SD ± 14.5) |
|
Sternorrhyncha |
249 |
793 |
409 |
255 |
129 |
155 |
264.3 (SD ± 140.2) |
|
Thysanoptera |
6 |
1977 |
951 |
776 |
250 |
320 |
659 (SD ± 364.9) |
|
Lepidoptera |
86 |
23 |
5 |
8 |
10 |
12 |
7.7 (SD ± 2.5) |
|
Non-target taxa of PT |
Acari |
14 |
206 |
34 |
53 |
119 |
170 |
68.7 (SD ± 44.6) |
Heteroptera |
15 |
70 |
22 |
41 |
7 |
7 |
23.3 (SD ± 17) |
|
Auchenorrhyncha |
74 |
63 |
23 |
19 |
21 |
24 |
21 (SD ± 2) |
|
Collembola |
28 |
32 |
3 |
17 |
12 |
12 |
10.7 (SD ± 7.1) |
|
Formicidae |
82 |
23 |
17 |
5 |
1 |
1 |
7.7 (SD ± 8.3) |
|
Araneae/Opiliones |
|
10 |
3 |
3 |
4 |
5 |
3.3 (SD ± 0.6) |
|
Mecoptera |
2 |
4 |
1 |
3 |
5 |
1.3 (SD ± 1.5) |
||
Dermaptera |
5 |
3 |
2 |
1 |
2 |
1 (SD ± 1) |
||
Psocoptera |
2 |
3 |
2 |
1 |
1 |
1 (SD ± 1) |
||
Orthoptera |
2 |
1 |
1 |
1 |
0.33 (SD ± 0.6) |
|||
Trichoptea |
2 |
|
|
|||||
Raphidioptera |
1 |
|
|
|||||
Total Abundance |
2289 |
12570 |
3366 |
4583 |
4621 |
5537 |
||
Abundance of target taxa (PT) |
12155 |
3261 |
4443 |
4451 |
5309 |
|||
Cerambycidae and Buprestidae
Within the order Coleoptera, 15 species of Cerambycidae were recorded with PT. An average of 9.7 ± 9 individuals and 5.7 ± 4 species per MAP field were assessed. White and yellow PT collected 55% and 45% of the individuals, respectively. A total of seven species of Buprestidae were collected, resulting in an average of 4 ± 1 species and 29.7 ± 21.1 individuals per MAP field. 90% of Buprestidae were collected with yellow PT and 10% with white PT. When PT B was considered, no differences in species richness for either family were recorded. The abundance of each species and species richness per MAP field is listed in Table 3.
Tab. 3: Abundance and species richness of Cerambycidae and Buprestidae recorded with Malaise traps (MT) in Merano/Meran (M) and with pan traps (PT) in each medicinal and aromatic plant field (M, K = Kastelruth/Castelrotto, W = Wiesen/Prati). For W, values with (+ PT B) and without (- PT B) the additional pan trap set are indicated.
+
Taxa |
PT |
MT |
||||
|---|---|---|---|---|---|---|
Cerambycidae |
Abundance |
M |
K |
W (- PT B) |
W (+ PT B) |
Abundance |
Pseudovadonia livida (Fabricius, 1776) |
3 |
|
|
3 |
3 |
1 |
Xylotrechus smei/stebbingi(Chevrolat, 1860) |
|
|
1 |
|||
Stenurella melanura (Linnaeus, 1758) |
8 |
3 |
2 |
2 |
|
|
Pachytodes cerambyciformis (Schrank, 1781) |
6 |
1 |
|
|
||
Cerambyx scopoli (Füssli, 1775) |
2 |
1 |
|
|
||
Dinoptera collaris (Linnaeus, 1758) |
7 |
6 |
10 |
|
||
Stenurella nigra (Linnaeus, 1758) |
7 |
1 |
2 |
|
||
Molorchus minor (Linnaeus, 1758) |
2 |
1 |
1 |
|
||
Gaurotes virginea (Linnaeus, 1758) |
3 |
2 |
2 |
|
||
Brachyta interrogationis (Linnaeus, 1758) |
3 |
1 |
1 |
|
||
Rudpela maculata (Poda, 1761) |
3 |
1 |
1 |
1 |
|
|
Chlorophorus figuratus (Scopoli, 1763) |
3 |
1 |
|
|
||
Chlorophorus sartor (O. F. Müller, 1766) |
2 |
1 |
|
|
||
Stenopterus ater (Linnaeus, 1767) |
2 |
1 |
|
|
||
Paracorymbiama culicornis (De Geer, 1775) |
3 |
2 |
2 |
|
||
Anastrangalia dubia (Scopoli, 1763) |
3 |
1 |
1 |
|
||
Abundance |
57 |
4 |
5 |
20 |
25 |
2 |
Species richness |
15 |
2 |
5 |
10 |
10 |
2 |
Buprestidae |
Abundance |
M |
K |
W (- PT B) |
W (+ PT B) |
|
Anthaxia quadripunctata (Linnaeus, 1758) |
71 |
10 |
42 |
19 |
19 |
|
Acmaeodera bipunctata (Olivier, 1790) |
4 |
1 |
3 |
|
||
Anthaxia podolica (Mannerheim, 1837) |
7 |
1 |
6 |
|
||
Anthaxia nitidula (Linnaeus, 1758) |
2 |
1 |
1 |
4 |
||
Anthaxia helvetica (Stierlin, 1868) |
3 |
3 |
4 |
|||
Agrilus integerrimus (Ratzeburg, 1837) |
1 |
1 |
|
|||
Agrilus viridis (Linnaeus, 1758) |
1 |
1 |
1 |
|||
Abundance |
89 |
12 |
53 |
24 |
28 |
|
Species richness |
7 |
3 |
5 |
4 |
4 |
|
Wild bees
Among the Hymenoptera, 235 individual wild bees were collected, and 47 different species identified in 10 genera (Fig. 3). One individual Andrena sp. and six Lasioglossum sp. were left at genus level. An average abundance of 78.3 ± 12.1 individuals and an average species richness of 27.7 ± 7.6 species per MAP field were recorded. The highest wild bee species richness was assessed in K with 36 species, followed by M with 26 and W with 21 species (Fig. 4). European Red List status [31] and functional traits (such as sociality, nesting type and pollen foraging type; [32][33]) of wild bees are shown in Figure 5. 78% of the collected oligolectic wild bees (host-plant specialist) are specialized on Asteraceae and 11% each on Caprifoliaceae and Campanulaceae [32][33]. A complete species list with abundances, species richness and average abundance of each species per MAP field, as well as the respective functional traits of wild bees, is found in Table 4 and Table 5, respectively.
×
- [31] Nieto A., Roberts S.P.M., Kemp J. et al. (2014). European Red List of bees. Publication Office of the European Union, Luxembourg, Luxembourg, DOI: 10.2779/77003.
- [32] Scheuchl E., Willner W. (2016). Taschenlexikon der Wildbienen. Alle Arten im Porträt. Quelle und Mayer, Wiebelsheim, Germany, here pp. 100-817.
- [33] Westrich P. (2018). Die Wildbienen Deutschlands. Ulmer, Stuttgart, Germany, here pp. 399-719.
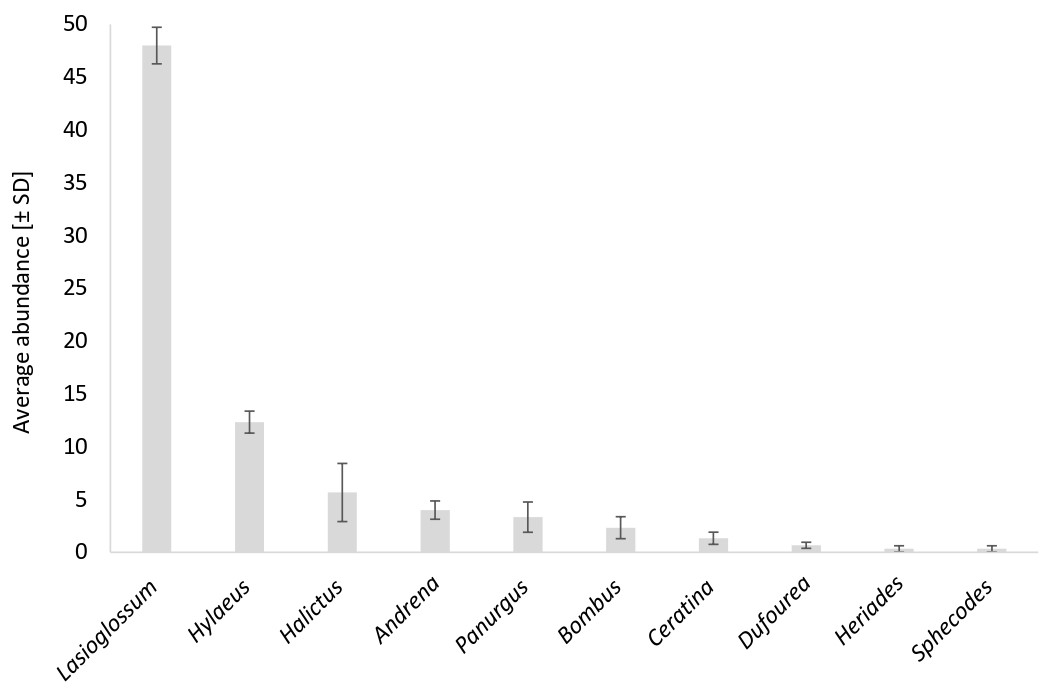
+
Fig. 3: Average abundance and standard deviation per medicinal and aromatic plant field for each wild bee genus, including individuals identified to genus level.
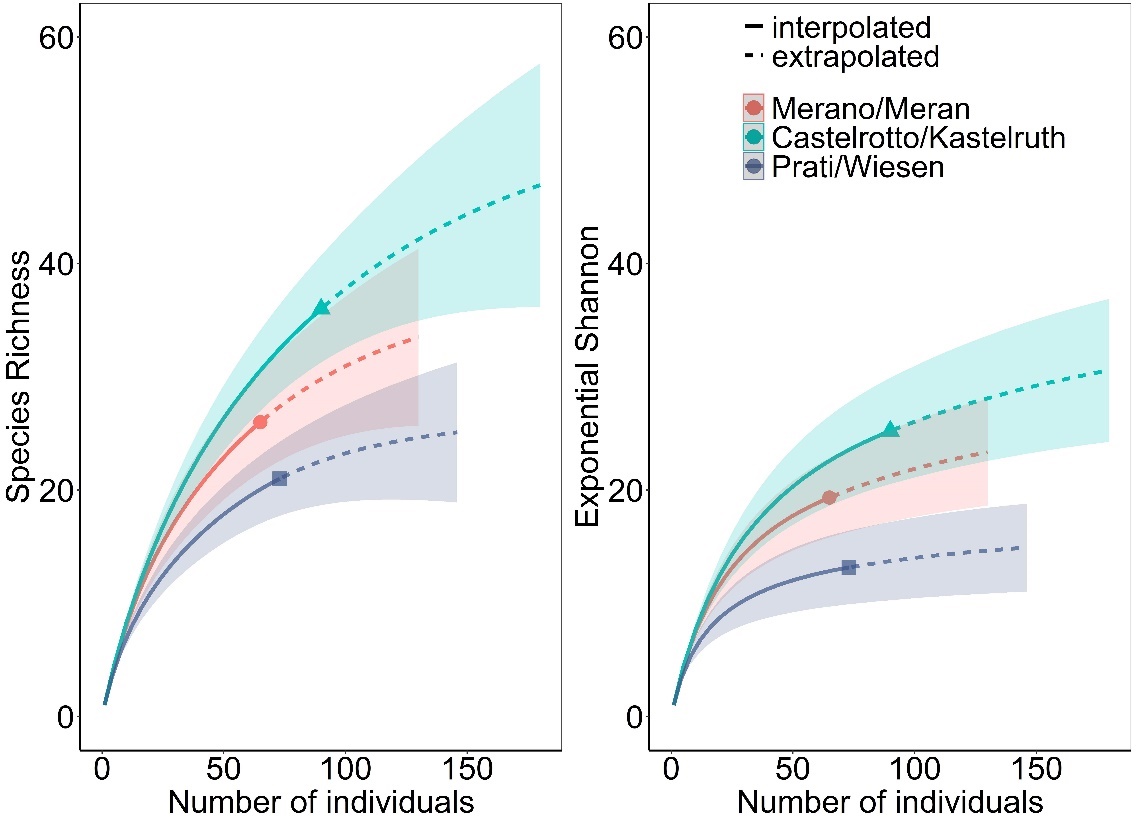
+
Fig. 4: Accumulation curves of wild bee species richness and wild bee diversity per medicinal and aromatic plant field. A higher wild bee diversity was recorded in Castelrotto/Kastelruth than in Prati/Wiesen, Merano/Meran scored intermediate.
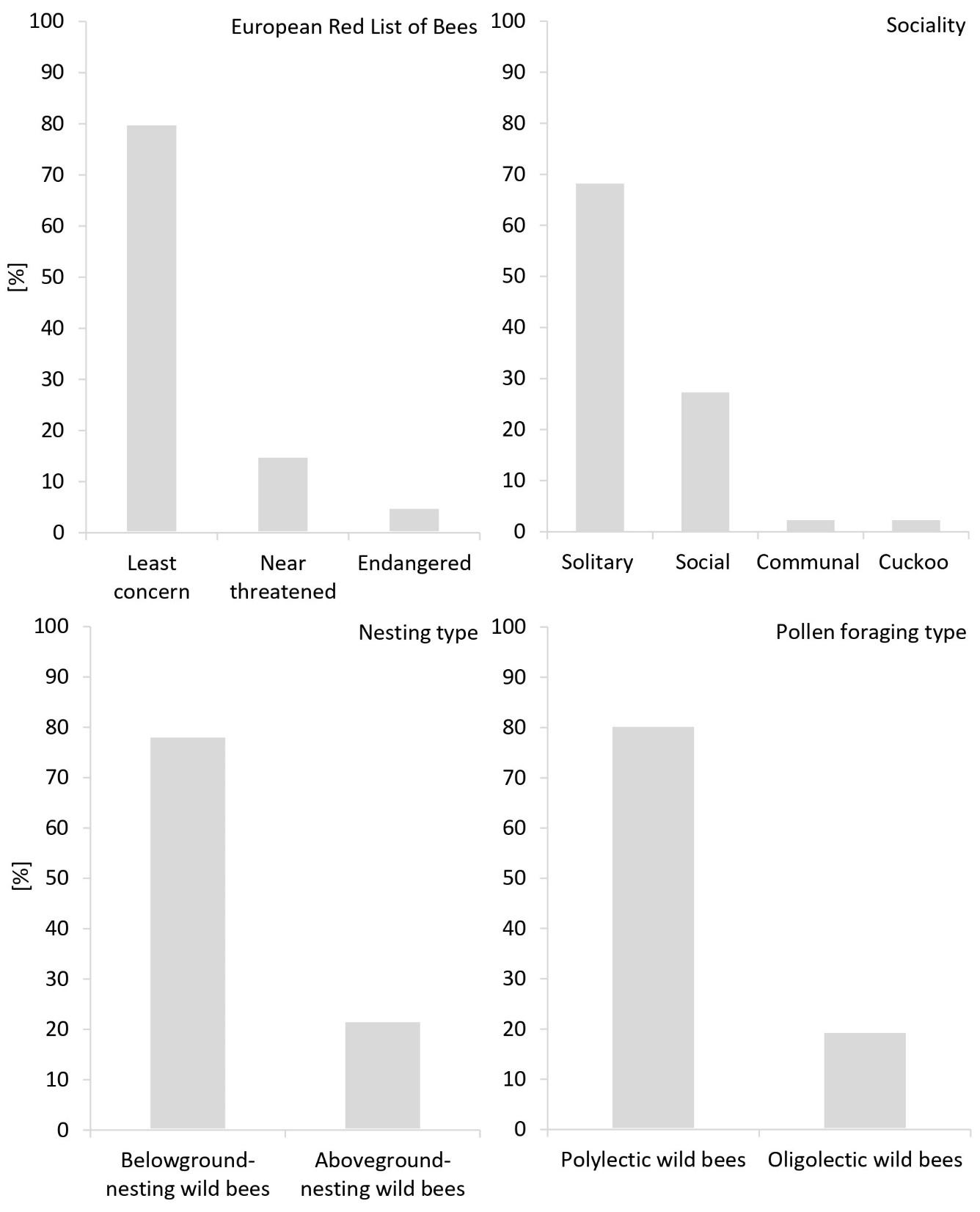
+
Fig. 5: Percentage of wild bees per European Red List category (46 sp.), sociality (47 sp.), nesting type (46 sp.) and pollen foraging type (46 sp.). For one wild bee speciesh, the European Red List category was not available. Cuckoo bees were excluded from nesting type and pollen foraging type.
Tab. 4: Abundance of wild bees recorded with Malaise traps (MT) in Merano/Meran (M) and with pan traps (PT) in each medicinal and aromatic plant field (MAP) (M, K = Kastelruth/Castelrotto, W = Wiesen/Prati). For W, values with (+ PT B) and without (- PT B) the additional pan trap set are indicated. Average abundance (± SD) per MAP field and species, which were collected with PT, were calculated with M, K, and W (- PT B).
+
MT |
PT |
||||||
|---|---|---|---|---|---|---|---|
Taxa |
Abundance |
Abundance |
M |
K |
W (-PT B) |
W (+ PT B) |
Average (± SD)/MAP field |
Andrena sp. |
|
1 |
|
|
1 |
1 |
0.3 (SD ± 0.6) |
Andrena fulvago (Christ, 1791) |
2 |
3 |
1 |
2 |
1 (SD ± 1) |
||
Andrena haemorrhoa (Fabricius, 1781) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Andrena hattorfiana (Fabricius, 1775) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Andrena humilis (Imhoff, 1832) |
|
2 |
2 |
2 |
0.7 (SD ± 1.2) |
||
Andrena minutula (Kirby, 1802) |
|
2 |
1 |
1 |
0.7 (SD ± 0.6) |
||
Andrena nitida (Müller, 1776) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Andrena ovatula (Kirby, 1802) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Bombus humilis (Illiger, 1806) |
1 |
|
|
||||
Bombus lapidarius (Linnaeus, 1758) |
1 |
6 |
3 |
3 |
2 (SD ± 1.7) |
||
Bombus lucorum (Linnaeus, 1761) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Ceratina cucurbitina (Rossi, 1792) |
|
4 |
2 |
2 |
1.3 (SD ± 1.2) |
||
Dufourea dentiventris (Nylander, 1848) |
|
1 |
1 |
1 |
0.3 (SD ± 0.6) |
||
Dufourea paradoxa (Morawitz, 1868) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Halictus eurygnathus (Blüthgen, 1931) |
|
2 |
1 |
1 |
0.7 (SD ± 0.6) |
||
Halictus maculatus (Smith, 1848) |
|
3 |
2 |
1 |
1 |
1 (SD ± 1) |
|
Halictus quadricinctus (Fabricius, 1776) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Halictus simplex (Blüthgen, 1923) |
|
6 |
6 |
2 (SD ± 3.5) |
|||
Halictus subauratus (Rossi, 1792) |
1 |
4 |
2 |
2 |
1.3 (SD ± 1.2) |
||
Halictus tumulorum (Linnaeus, 1758) |
1 |
1 |
1 |
1 |
0.3 (SD ± 0.6) |
||
Heriades crenulatus (Nylander, 1856) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Hylaeus annularis (Kirby, 1802) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Hylaeus annulatus (Linnaeus, 1758) |
|
2 |
1 |
1 |
1 |
0.7 (SD ± 0.6) |
|
Hylaeus brevicornis (Nylander, 1852) |
|
4 |
2 |
2 |
3 |
1.3 (SD ± 1.2) |
|
Hylaeus communis (Nylander, 1852) |
|
10 |
2 |
3 |
5 |
5 |
3.3 (SD ± 1.5) |
Hylaeus confusus (Nylander, 1852) |
|
12 |
4 |
6 |
2 |
2 |
4 (SD ± 2) |
Hylaeus hyalinatus (Smith, 1842) |
|
14 |
6 |
8 |
8 |
4.7 (SD ± 4.2) |
|
Hylaeus nigritus (Fabricius, 1798) |
|
3 |
1 |
1 |
1 |
1 |
1 (SD ± 0) |
Lasioglossum sp. |
|
6 |
2 |
1 |
3 |
3 |
2 (SD ± 1) |
Lasioglossum albipes (Fabricius, 1781) |
2 |
2 |
1 |
1 |
1 |
0.7 (SD ± 0.6) |
|
Lasioglossum calceatum (Scopoli, 1763) |
2 |
14 |
5 |
2 |
7 |
7 |
4.7 (SD ± 2.5) |
Lasioglossum corvinum (Morawitz, 1877) |
|
3 |
2 |
1 |
1 (SD ± 1) |
||
Lasioglossum griseolum (Morawitz, 1872) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Lasioglossum interruptum (Panzer, 1798) |
|
4 |
1 |
3 |
1.3 (SD ± 1.5) |
||
Lasioglossum laeve (Kirby, 1802) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Lasioglossum laevigatum (Kirby, 1802) |
1 |
29 |
7 |
3 |
19 |
19 |
9.7 (SD ± 8.3) |
Lasioglossum leucopus (Kirby, 1802) |
|
4 |
1 |
1 |
2 |
2 |
1.3 (SD ± 0.6) |
Lasioglossum lucidulum (Schenk, 1861) |
|
3 |
3 |
1 (SD ± 1.7) |
|||
Lasioglossum minutulum (Schenk, 1853) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Lasioglossum morio (Fabricius, 1793) |
3 |
27 |
9 |
15 |
3 |
4 |
9 (SD ± 6) |
Lasioglossum pauperatum (Brullé, 1832) |
|
2 |
2 |
0.7 (SD ± 1.2) |
|||
Lasioglossum planulum (Pérez, 1903) |
1 |
|
|
||||
Lasioglossum politum (Schenk, 1853) |
|
6 |
6 |
2 (SD ± 3.5) |
|||
Lasioglossum punctatissimum (Schenk, 1853) |
3 |
|
|
||||
Lasioglossum quadrisignatum (Schenk, 1853) |
|
3 |
1 |
2 |
2 |
1 (SD ± 1) |
|
Lasioglossum semilucens (Alfken, 1914) |
2 |
5 |
3 |
2 |
2 |
1.7 (SD ± 1.5) |
|
Lasioglossum subhirtum (Lepeletier, 1841) |
|
6 |
2 |
3 |
1 |
1 |
2 (SD ± 1) |
Lasioglossum tricinctum (Schenk, 1874) |
|
4 |
1 |
2 |
1 |
2 |
1.3(SD ± 0.6) |
Lasioglossum zonulum (Smith, 1848) |
|
14 |
7 |
3 |
4 |
4 |
4.7 (SD ± 2.1) |
Panurgus banksianus (Kirby, 1802) |
|
8 |
1 |
7 |
8 |
2.7 (SD ± 3.8) |
|
Panurgus calcaratus (Scopoli, 1763) |
|
2 |
2 |
0.7 (SD ± 1.2) |
|||
Sphecodes niger (Hagens, 1874) |
|
1 |
1 |
0.3 (SD ± 0.6) |
|||
Abundance |
20 |
235 |
67 |
91 |
77 |
81 |
|
Species Richness |
12 |
47 |
26 |
36 |
21 |
21 |
|
Tab. 5: Red List category (EN = endangered, NT = near threatened, LC = least concern, DD = data deficient) and functional traits (PFT = pollen foraging type, PFS = plant family specialization, S = Sociality, NT = Nesting type) of wild bees.
+
Taxa |
Red List [31]
×
|
PFT |
PFS |
S |
NT |
|---|---|---|---|---|---|
Andrena fulvago |
DD |
oligolectic |
Asteraceae |
solitary |
below-ground |
Andrena haemorrhoa |
LC |
polylectic |
solitary |
below-ground |
|
Andrena hattorfiana |
NT |
oligolectic |
Caprifoliaceae |
solitary |
below-ground |
Andrena humilis |
DD |
oligolectic |
Asteraceae |
solitary |
below-ground |
Andrena minutula |
DD |
polylectic |
solitary |
below-ground |
|
Andrena nitida |
LC |
polylectic |
solitary |
below-ground |
|
Andrena ovatula |
NT |
polylectic |
solitary |
below-ground |
|
Bombus humilis |
LC |
polylectic |
eusocial |
above-ground |
|
Bombus lapidarius |
LC |
polylectic |
eusocial |
above-ground |
|
Bombus lucorum |
LC |
polylectic |
eusocial |
below-ground |
|
Ceratina cucurbitina |
LC |
polylectic |
solitary |
above-ground |
|
Dufourea dentiventris |
NT |
oligolectic |
Campanulaceae |
solitary |
below-ground |
Dufourea paradoxa |
LC |
polylectic |
solitary |
below-ground |
|
Halictus eurygnathus |
|
polylectic |
solitary |
below-ground |
|
Halictus maculatus |
LC |
polylectic |
eusocial |
below-ground |
|
Halictus quadricinctus |
NT |
polylectic |
solitary |
below-ground |
|
Halictus simplex |
LC |
polylectic |
solitary |
below-ground |
|
Halictus subauratus |
LC |
polylectic |
eusocial |
below-ground |
|
Halictus tumulorum |
LC |
polylectic |
eusocial |
below-ground |
|
Heriades crenulatus |
LC |
oligolectic |
Asteraceae |
solitary |
above-ground |
Hylaeus annularis |
DD |
oligolectic |
Asteraceae |
solitary |
above-ground |
Hylaeus annulatus |
DD |
polylectic |
solitary |
above-ground |
|
Hylaeus brevicornis |
LC |
polylectic |
solitary |
above-ground |
|
Hylaeus communis |
LC |
polylectic |
solitary |
above-ground |
|
Hylaeus confusus |
LC |
polylectic |
solitary |
above-ground |
|
Hylaeus hyalinatus |
LC |
polylectic |
solitary |
above-ground |
|
Hylaeus nigritus |
LC |
oligolectic |
Asteraceae |
solitary |
above-ground |
Lasioglossum albipes |
LC |
polylectic |
eusocial |
below-ground |
|
Lasioglossum calceatum |
LC |
polylectic |
eusocial |
below-ground |
|
Lasioglossum corvinum |
LC |
polylectic |
below-ground |
||
Lasioglossum griseolum |
LC |
polylectic |
below-ground |
||
Lasioglossum interruptum |
LC |
polylectic |
eusocial |
below-ground |
|
Lasioglossum laeve |
EN |
polylectic |
solitary |
below-ground |
|
Lasioglossum laevigatum |
NT |
polylectic |
solitary |
below-ground |
|
Lasioglossum leucopus |
LC |
polylectic |
solitary |
below-ground |
|
Lasioglossum lucidulum |
LC |
polylectic |
solitary |
below-ground |
|
Lasioglossum minutulum |
NT |
polylectic |
solitary |
below-ground |
|
Lasioglossum morio |
LC |
polylectic |
eusocial |
below-ground |
|
Lasioglossum pauperatum |
LC |
polylectic |
below-ground |
||
Lasioglossum planulum |
|
polylectic |
below-ground |
||
Lasioglossum politum |
LC |
polylectic |
eusocial |
below-ground |
|
Lasioglossum punctatissimum |
LC |
polylectic |
below-ground |
||
Lasioglossum quadrisignatum |
EN |
polylectic |
below-ground |
||
Lasioglossum semilucens |
LC |
polylectic |
solitary |
below-ground |
|
Lasioglossum subhirtum |
LC |
polylectic |
below-ground |
||
Lasioglossum tricinctum |
DD |
polylectic |
eusocial |
below-ground |
|
Lasioglossum zonulum |
LC |
polylectic |
solitary |
below-ground |
|
Panurgus banksianus |
LC |
oligolectic |
Asteraceae |
solitary |
below-ground |
Panurgus calcaratus |
LC |
oligolectic |
Asteraceae |
communal |
below-ground |
Sphecodes niger |
LC |
|
|
cuckoo bee of Lasioglossum morio |
|
Flowering MAPs and pollinators
Abundance of Syrphidae, A. mellifera and wildbees did not correlate significantly with MAP flower abundance. However, a positive correlation was detected for wild bee species richness (Fig. 6).
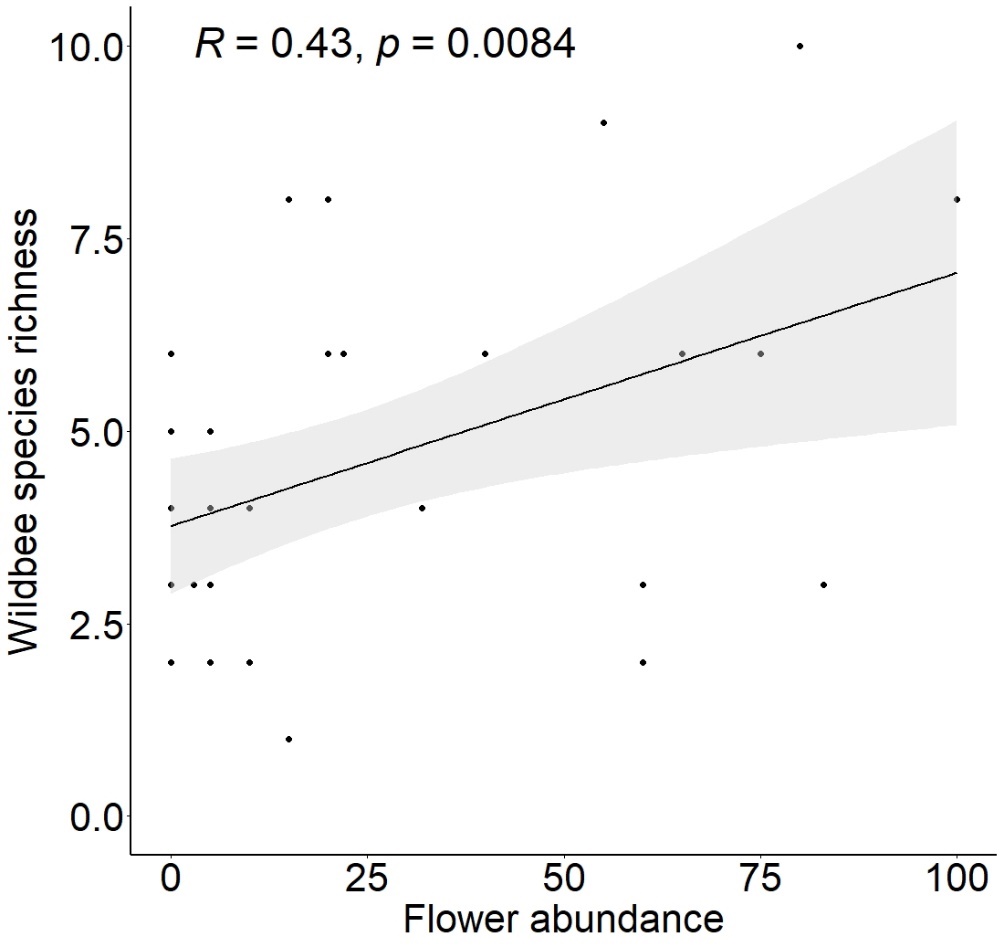
+
Fig. 6: Significant effect of flower abundance on wild bee species richness. Flower abundance was calculated as the percentage of medicinal and aromatic plants in flower (0-100) around each pan trap set. Dots represent raw data per survey event and bands represent the 95% confidence interval.
Hymenopteran parasitoids
A total of 334 parasitoids from the Hymenoptera order were detected, belonging to 14 parasitoid families and two superfamilies. On average of 111.3 ± 34 parasitoids per MAP field were recorded. Aphelinidae, Encyrtidae and Aphidiinae (Braconidae), which are mostly parasitoids of Sternorrhyncha [22], accounted for 38.4% of the total parasitoids in PT. Abundance of each hymenopteran parasitoid family/superfamily per MAP field is listed in Table 6.
×
- [22] Goulet H., Huber J.T. (eds.) (1993). Hymenoptera of the world. An identification guide to families. Canada Communication Group, Ottawa, Canada, here pp. 65-529.
Tab. 6: Abundance and family richness of Hymenopteran parasitoids recorded with Malaise traps in Merano/Meran (M) and with pan traps in each medicinal and aromatic plant field (MAP) (M, K = Kastelruth/Castelrotto, W = Wiesen/Prati). For W, values with (+ PT B) and without (- PT B) the additional pan trap set are indicated. Average abundance (± SD) per MAP field and taxon were calculated with M, K, and W (- PT B).
+
MT |
PT |
||||||
|---|---|---|---|---|---|---|---|
Taxa |
Abundance |
Abundance |
M |
K |
W (-PT B) |
W (+ PT B) |
Average (± SD)/MAP field |
Braconidae varia |
68 |
30 |
7 |
14 |
9 |
9 |
10 (SD ± 3.6) |
Aphidiinae (Braconidae) |
31 |
11 |
1 |
9 |
1 |
1 |
3.7 (SD ± 4.6) |
Ichneumonidae |
33 |
10 |
6 |
1 |
3 |
3 |
3.3 (SD ± 2.5) |
Cynipoidea varia |
64 |
19 |
4 |
5 |
10 |
12 |
6.3 (SD ± 3.2) |
Chalcidoidea varia (mostly Pteromalidae/Tetracampidae) |
23 |
13 |
3 |
3 |
7 |
7 |
4.3 (SD ± 2.3) |
Aphelinidae |
20 |
9 |
4 |
3 |
2 |
3 |
3 (SD ± 1) |
Chalcididae |
1 |
|
|
||||
Elasmidae |
1 |
|
|
||||
Encyrtidae |
29 |
101 |
53 |
48 |
1 |
50.5 (SD ± 3.5) |
|
Eulophidae |
16 |
16 |
6 |
5 |
5 |
7 |
5.3 (SD ± 0.6) |
Eurytomidae |
2 |
2 |
2 |
2 |
|||
Mymaridae |
120 |
21 |
4 |
11 |
6 |
10 |
7 (SD ± 3.6) |
Signiphoridae |
2 |
|
|
||||
Trichogrammatidae |
4 |
2 |
1 |
1 |
1 |
1 (SD ± 0) |
|
Torymidae |
1 |
|
|
||||
Ceraphronidae |
7 |
19 |
3 |
3 |
13 |
14 |
6.3 (SD ± 5.8) |
Megaspilidae |
2 |
|
|
||||
Proctotrupidae |
10 |
|
|
||||
Diapriidae |
24 |
13 |
3 |
9 |
1 |
1 |
4.3 (SD ± 4.2) |
Evaniidae |
2 |
|
|
||||
Platygastridae |
19 |
11 |
2 |
5 |
4 |
6 |
3.7 (SD ± 1.5) |
Scelionidae |
26 |
52 |
27 |
16 |
9 |
13 |
17.3 (SD ± 9.1) |
Chrysididae |
3 |
5 |
3 |
2 |
2 |
2.5 (SD ± 0.7) |
|
Dryinidae |
3 |
|
|
||||
Bethylidae |
3 |
|
|
||||
Abundance |
514 |
334 |
123 |
138 |
73 |
90 |
|
Family richness (superfamilies excluded) |
22 |
13 |
9 |
12 |
10 |
11 |
|
Additional surveys
Malaise traps
Overall, 2289 arthropods were collected with Malaise traps, resulting in an average of 763 ± 338.9 individuals recorded per survey event, with the same taxa recorded as with PT. The exceptions were Buprestidae and Araneae/Opiliones, which were not detected with Malaise traps, and Raphidioptera and Trichoptera, which were only observed in Malaise traps. A complete list with total abundances is found in Table 2. One of the two species of Cerambycidae was only recorded with Malaise traps. Therefore, a total of 16 Cerambycidae species were found in MAP fields when both Malaise traps and PT were combined (Table 3). A total of 20 wild bees were recorded with Malaise traps. Of the 12 wild bee species collected, three were previously not detected with PT (Table 4). This results in a total of 50 wild bee species, which corresponds to 10% of the estimated South Tyrolean wild bee fauna [34]. A total of 514 hymenopteran parasitoids were collected, of which 16% were parasitoids of Sternorrhyncha. Nine of the 22 families detected with Malaise traps were observed exclusively in Malaise traps (Tab. 6).
×
- [34] Hellrigl K. (2003). Faunistik der Ameisen und Wildbienen Südtirols (Hymenoptera: Formicidae et Apoidea). Gredleriana 3, 143-208.
Grasshoppers, butterflies, birds, and bats
Nine grasshoppers and 117 butterflies were counted in M, which were assigned to five and eight species, respectively. A total of 36 and 35 birds were counted in M and W, respectively, which were assigned to 15 and 14 bird species. Ten species of bats were recorded in M, five of which were recorded while foraging. Bat genera were counted as distinct species, with the exception of Pipistrellus sp.. The specimen recorded could not be assigned to one of three Pipistrellus species assessed and was therefore not counted as distinct species. An additional descriptive analysis regarding the functional guilds of bats was performed. Total calls of each species/genus were assigned to the bats functional guilds [35], resulting in 55% narrow space foragers, 49% edge space foragers and 18% open space foragers. Of the six bat species detected foraging, four were edge space foragers, one a narrow space forager and one an open space forager.
×
- [35] Huemer P. (2004). Die Tagfalter Südtirols (Veröffentlichungen des Naturmuseums Suedtirol , 2). Folio Verlag, Vienna, Austria.
Abundance of each species and their respective Red List categories [36][37][38][39] are listed in Table 7 and Table 8.
×
- [36] Ceresa F., Kranebitter P. (2020). Lista Rossa 2020 degli uccelli nidificanti in Alto Adige. Gredleriana 20, 57-70.
- [37] Hilpold A., Wilhalm T., Kranebitter P. (2017). Rote Liste der gefährdeten Fang- und Heuschrecken Südtirols (Insecta: Orthoptera, Mantodea). Gredleriana 17, 61-86.
- [38] Rondinini C., Battistoni A., Teofili C. (2022). Lista Rossa IUCN dei Vertebrati Italiani 2022. Comitato Italiano IUCN e Ministero dell'Ambiente e della Tutela del Territorio e del Mare, Roma, Italia, here p. 31.
- [39] Dietz C., Kiefer A. (2016). Bats of Britain and Europe. Bloomsbury Publishing, London, United Kingdom, here pp. 104-105.
Tab. 7: Abundance and Red List category (LC = least concern, NE = not evaluated, NT = near threatened) of grasshopper and butterfly species recorded in M (Merano/Meran).
+
Grasshoppers |
Abundance |
|
Chorthippus brunneus (Linnaeus, 1758) |
4 |
LC |
Mantis religiosa (Linnaeus, 1758) |
2 |
LC |
Gomphocerippus rufus (Linnaeus, 1758) |
1 |
LC |
Gryllus campestris (Linnaeus, 1758) |
1 |
LC |
Oedipoda caerulescens (Linnaeus, 1758) |
1 |
LC |
Butterflies |
Abundance |
|
Pieris rapae (Linnaeus, 1758) |
79 |
LC |
Polyommatus icarus (Rottemburg, 1775) |
23 |
LC |
Pieris napi (Linnaeus, 1758) |
6 |
LC |
Vanessa cardui (Linnaeus, 1758) |
3 |
NE |
Colias croceus (Geoffrey in Fourcroy, 1785) |
2 |
NE |
Vanessa atalanta (Linnaeus, 1758) |
2 |
NE |
Lysandra bellargus (Rottemburg, 1775) |
1 |
LC |
Pieris brassicae (Linnaeus, 1758) |
1 |
NT |
Tab. 8: Abundance and Red List category (EN = endangered, VU = vulnerable, NT = near threatened, LC = least concern, DD = data deficient, NE = not evaluated) of birds and bats in medicinal and aromatic plant fields. For birds, abundance in M (Meran/Merano) and W (Wiesen/Prati) are listed separately. For bats recorded in M, the total calls and the number of feeding buzzes are listed.
+
Birds |
Abundance M |
Abundance W |
|
|---|---|---|---|
Passer italiae (Vieillot, 1817) |
|
10 |
VU |
Turdus merula (Linnaeus, 1758) |
7 |
3 |
LC |
Sylvia atricapilla (Linnaeus, 1758) |
5 |
4 |
LC |
Passer montanus (Linnaeus, 1758) |
5 |
1 |
EN |
Hirundo rustica (Linnaeus, 1758) |
5 |
LC |
|
Carduelis carduelis (Linnaeus, 1758) |
4 |
|
LC |
Corvus corone (Linnaeus, 1758) |
1 |
2 |
LC |
Fringilla coelebs (Linnaeus, 1758) |
3 |
|
LC |
Serinus serinus (Linnaeus, 1766) |
2 |
1 |
LC |
Chloris chloris (Linnaeus, 1758) |
1 |
1 |
EN |
Parus major (Linnaeus, 1758) |
2 |
LC |
|
Phoenicurus ochruros (S. G. Gmelin, 1774) |
2 |
LC |
|
Pica pica (Linnaeus, 1758) |
1 |
1 |
LC |
Turdus pilaris (Linnaeus, 1758) |
2 |
|
NT |
Aquila chrysaetos (Linnaeus, 1758) |
1 |
VU |
|
Columba palumbus (Linnaeus, 1758) |
1 |
|
LC |
Dendrocopos major (Linnaeus, 1758) |
1 |
|
LC |
Erithacus rubecula (Linnaeus, 1758) |
1 |
LC |
|
Jynx torquilla (Linnaeus, 1758) |
1 |
|
DD |
Phoenicurus phoenicurus (Linnaeus, 1758) |
1 |
VU |
|
Turdus philomelos (C. L. Brehm, 1831) |
1 |
|
LC |
Turdus viscivorus (Linnaeus, 1758) |
1 |
|
LC |
Bats |
Total calls |
Feeding buzzes |
|
Rhinolophus hipposideros (Borkhausen, 1797) |
38 |
1 |
EN |
Pipistrellus pipistrellus (Schreber, 1774) |
27 |
6 |
LC |
Nyctalus/Vespertilio/Eptesicus |
17 |
1 |
|
Pipistrellus kuhlii/nathusii |
10 |
4 |
LC/NT |
Myotis myotis/Myotis blythii |
9 |
|
VU |
Pipistrellus pygmaeus (Leach, 1825) |
9 |
2 |
NT |
Myotis sp. |
7 |
|
|
Hypsugo savii (Bonaparte, 1837) |
2 |
1 |
LC |
Nyctalus leisleri (Kuhl, 1817) |
1 |
|
NT |
Pipistrellus sp. |
1 |
|
|
Plecotus macrobullaris/auritus |
1 |
|
EN/NT |
Ground-dwelling macro-invertebrates
Pitfall traps collected a total of 352 individuals, of which 254 and 98 individuals were classified as target and non-target taxa, respectively. Formicidae (n = 91), Staphylinidae (n = 34), Carabidae (n = 33), and Araneae (n = 29) were the most abundant taxa, accounting for 74% of all target taxa. For descriptive analysis, target taxa were categorized according to their respective trophic guilds [40][41][42][43][44][45][46] (Fig. 7). Trophic guild assignment and respective abundances are in Table 9.
×
- [40] Origazzi A., Bardgett R., Barrios E. et al. (2016). Global Soil Biodiversity Atlas. Publication Office of the European Union, Luxembourg, Luxembourg, here pp. 28-65.
- [41] Bellmann H. (20063). Kosmos-Atlas Spinnentiere Europas. Kosmos, Stuttgart, Germany, here pp. 20-254.
- [42] Koch K. (1999). Carabidae - Micropeplidae. In: Die Käfer Mitteleuropas (Bd. E1). Spektrum Akademischer Verlag, Heidelberg, Germany.
- [43] Schöller M. (1996). Ökologie mitteleuropäischer Blattkäfer, Samenkäfer und Breitrüssler. Eigenverlag des EVCV, Bürs, Austria, here pp. 5-11.
- [44] Rheinheimer J., Hassler M. (2013). Rüsselkäfer Baden-Württembergs. Verlag Regionalkultur, Heidelberg, Germany, here pp. 31-67.
- [45] Klausnitzer B. (2011). Stresemann - Exkursionsfauna von Deutschland. Bd. 2: Wirbellose. Insekten. Spektrum Akademischer Verlag, Heidelberg, Germany, here pp. 111-316.
- [46] Harde K.W., Helb M., Elzner K. (2021). Der Kosmos Käferführer. Franckh-Kosmos Verlag, Stuttgart, Germany, here pp. 96-259.
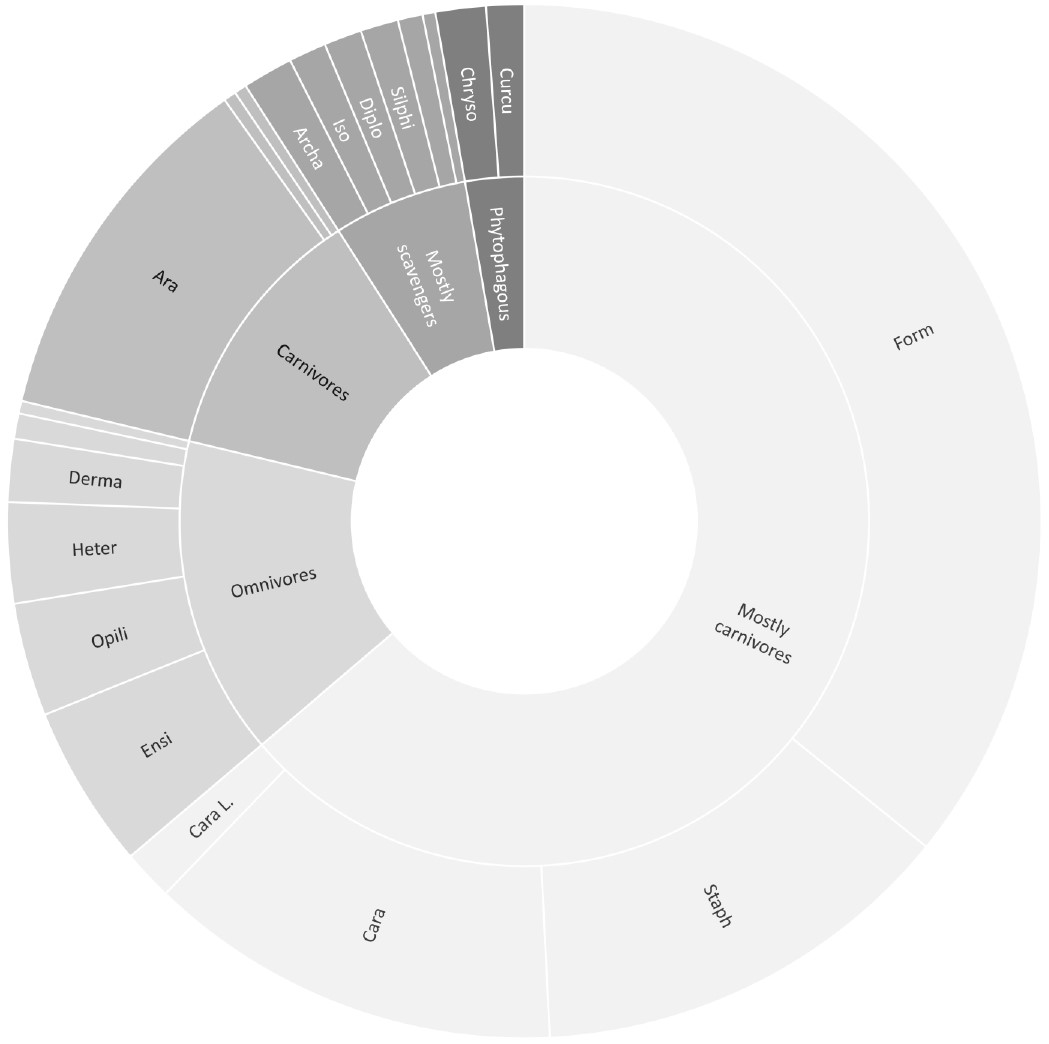
+
Fig. 7: Percentage of taxa collected with pitfall traps, subdivided according to their trophic guild. Only target taxa of pitfall traps were included (Form = Formicidae, Staph = Staphilinidae, Cara = Carabidae, Cara L. = Carabidae Larvae Ensi = Ensifera, Opili = Opiliones, Heter = Heteroptera, Derma = Dermaptera, Ara = Araneae, Archa = Archaeognatha, Iso = Isopoda, Diplo = Diplopoda, Silphi = Silphidae, Chryso = Chrysomelidae, Curcu = Curculionidae).
Tab. 9: Abundance of ground-dwelling macro-invertebrates collected with pitfall traps in Merano/Meran. Ground-dwelling macro-invertebrates are divided into non-target and target taxa. For the latter, the trophic guild and corresponding reference are listed.
+
Taxa |
Abundance |
Trophic guild |
Reference |
|
|---|---|---|---|---|
Target taxa |
Isopoda |
3 |
Scavengers (mostly) |
|
Araneae |
29 |
Carnivors |
||
Pseudoscorpiones |
1 |
Carnivors |
||
Opiliones |
9 |
Omnivores |
||
Chilopoda_Lithobiidae |
1 |
Carnivors |
||
Diplopoda_Julidae |
3 |
Scavengers (mostly) |
||
Carabidae |
33 |
Carnivors (mostly) |
||
Chrysomelidae |
4 |
Phytophagous |
||
Curculionidae |
3 |
Phytophagous |
||
Elateridae |
1 |
Omnivores |
||
Scarabaeidae |
2 |
Omnivores |
||
Silphidae |
3 |
Scavengers (mostly) |
||
Staphylinidae |
34 |
Carnivors (mostly) |
||
Carabidae Larvae |
4 |
Carnivors (mostly) |
||
Silphidae Larvae |
2 |
Scavengers (mostly) |
||
Dermestidae Larvae |
1 |
Scavengers (mostly) |
||
Formicidae |
91 |
Carnivors (mostly) |
||
Heteroptera |
8 |
Omnivores |
||
Dermaptera |
5 |
Omnivores |
||
Ensifera |
13 |
Omnivores |
||
Archaeognatha |
4 |
Scavengers (mostly) |
||
Non-target taxa |
Lumbricidae |
2 |
||
Brachycera/Nematocera |
65 |
|||
Hymenoptera |
8 |
|||
Sternorrhyncha |
1 |
|||
Auchenorrhyncha |
21 |
|||
Lepidoptera Larvae |
1 |
Discussion
In the following descriptive study, medicinal and aromatic plant (MAP) fields were surveyed from a multi-taxonomic perspective by taking the invertebrate and vertebrate fauna into account. MAP fields in South Tyrol represent small, structurally diverse and complex farmlands, rich in non-crop elements (e.g., dry-stone walls, hedges, etc.) and with a high diversity of plants grown on a relatively small area. High levels of habitat heterogeneity in agricultural areas are positively related to the suitability of farmlands as breeding and foraging habitats for several taxa [10][47]. The suitability of MAP fields as such is reflected in the high abundance and diversity of arthropods and vertebrates in our survey. Flower-visiting insects, in particular, appear to have found a valuable habitat in MAP fields, as indicated by the high number of pollinators and other insects captured with flower-mimicking pan traps (PT). Buprestidae were clearly attracted to the color yellow (PT) (as also noted in [48][49]), while Cerambycidae were attracted to both white and yellow PT [48]. Some genera (e.g., Buprestidae: Anthaxia sp.) or subfamilies (e.g., Cerambycidae: Lepturinae) within these two families feed primarily on pollen in the adult stage. Anthophagous Cerambycidae, for example, prefer Apiaceae and Asteraceae [50] and are found on flower heads with easily accessible pollen [48], as found in MAP fields. Among the pollinators, wild bees were by far the most abundant group, followed by Syrphidae and managed Apis mellifera.
×
- [10] Licata M., Maggio A.M., La Bella S. et al. (2022). Medicinal and aromatic plants in agricultural research, when considering criteria of multifunctionality and sustainability. Agriculture 12 (4), 529, DOI: 10.3390/agriculture12040529.
- [47] Wilson J.D., Whittingham M.J., Bradbury R.D. (2005). The management of crop structure. A general approach to reversing the impacts of agricultural intensification on birds? Ibis 147 (3), 453-463, DOI: 10.1111/j.1474-919x.2005.00440.x.
- [48] O’Neill K.M., Fultz J.E., Ivie M.A. (2008). Distribution of adult Cerambycidae and Buprestidae (Coleoptera) in a subalpine forest under shelterwood management. The Coleopterists Bulletin, 62 (1), 27-36.
- [49] Sakalian V., Langourov M. (2004). Colour traps a method for distributional and ecological investigations of Buprestidae (Coleoptera). Acta Societatis Zoologicae Bohemicae 68, 53-59.
- [50] Klausnitzer B., Klausnitzer U., Wachmann E. et al. (2016). Die Bockkäfer Mitteleuropas. VerlagsKG Wolf, Magdeburg, Germany, here pp. 118-133.
This was unexpected, as most studies dealing with pollinators in agroecosystems report higher abundances of A. mellifera than wild bees [51][52]. To properly understand this pattern, further information regarding the distribution of A. mellifera-hives would be needed. However, considering that 10% of the regional wild bee species pool has been documented in only three MAP fields and that wild bee species richness positively correlated with flower cover, it appears that this habitat type is particularly suitable and valuable for wild bees. Lamiaceae, which was the predominant MAP family in the surroundings of the traps, represent a very important pollen source for wild bees and insects in general [17][53]. In addition, recent findings suggest that several taxa within Asteraceae (e.g., Centaurea sp., Anthemis sp., etc.), the second most abundant plant family, are crucial host plants for oligolectic wild bees [54]. Since most of the detected oligolectic wild bees were in fact specialized on Asteraceae, we assume that this habitat and the respective cultivated MAPs are particularly favorable for these species. Wild bees specialized on Campanulaceae and Caprifoliaceae, neither of which were cultivated, indicating the influence of the surrounding landscape on the wild bee diversity of MAP fields. In fact, wild bees are central-place foragers that forage only a few hundred meters around their nesting sites and are therefore particularly dependent on the nearby landscape [51]. In addition, small-scale MAP fields in our study may have served as ideal nesting sites, as MAP fields were rich in non-crop elements, such as dry-stone walls and hedges [55].
×
- [51] Földesi R., Kovács-Hostyánszki A., Korösi A. et al. (2016). Relationships between wild bees, hoverflies and pollination success in apple orchards with different landscape contexts. Agricultural and Forest Entomology 18 (1), 68-75, DOI: 10.1111/afe.12135.
- [52] Roquer-Beni L., Alins G., Arnan X., et al. (2021). Management-dependent effects of pollinator functional diversity on apple pollination services: A response-effect trait approach. Journal of Applied Ecology 58 (12), 2843-2853, DOI: 10.1111/1365-2664.14022.
- [17] Venkatesh Y.N., Neethu T., Ashajyothi M. et al. (2022). Pollinator activity and their role on seed set of medicinal and aromatic Lamiaceae plants. Journal of Apicultural Research, Vol. ahead-of-print, DOI: 10.1080/00218839.2022.2080949.
- [53] Ahrné K., Bengtsson J., Elmqvist T. (2009). Bumble Bees (Bombus spp) along a gradient of increasing urbanization. PLoS One 4:e5574, DOI: 10.1371/journal.pone.0005574.
- [54] Kuppler J., Neumüller U, Mayr A.V. et al. (2023). Favourite plants of wild bees. Agriculture, Ecosystems and Environment 342:108266, DOI: 10.1016/j.agee.2022.108266.
- [55] Schindler M., Diestelhorst O., Härtel S. et al. (2013). Monitoring agricultural ecosystems by using wild bees as environmental indicators. BioRisk 8, 53-71, DOI: 10.3897/biorisk.8.3600.
Among the arthropod taxa found in MAP fields, not only beneficial arthropods, such as pollinators, but also numerous potential pests were found. Sternorrhyncha were the most abundant, which may be due in part to this taxon’s high attraction to the trap colors [14][56]. However, ground-dwelling predators and parasitoids were also highly abundant and diversified. Particularly numerous were parasitoids of Sternorrhyncha, which are efficient and often host-specific natural enemies commonly used in biological control programs [57]. Considering that several studies have assessed the positive effect of high plant diversity on natural enemies and food webs [58][59][60], a high stability of arthropod communities across trophic levels might be assumed to occur in the surveyed MAP fields.
×
- [14] Meyer U., Blum H., Gärber U. et al. (2010). Praxisleitfaden Krankheiten und Schädlinge im Arznei- und Gewürzpflanzenanbau. Julius-Kühn-Institut Selbstverlag, Braunschweig, Germany.
- [56] Höring T.F. (2014). How aphids find their host plants, and how they don’t. Annals of Applied Biology 165 (1), 3-26, DOI: 10.1111/aab.12142.
- [57] Singh R., Singh G. (2016). Aphids and their biocontrol. In: Omkar (ed.). Ecofriendly Pest Management for Food Security. Academic Press, Cambridge, Massachusetts, USA, here pp. 63-108.
- [58] Haddad N.M., Crutsinger G.M., Gross K. et al. (2010). Plant diversity and the stability of food webs. Ecology Letters 14 (1), 42-46, DOI: 10.1111/j.1461-0248.2010.01548.x.
- [59] Wan N.-F., Zheng X.-R., Fu L.-W. et al. (2020). Global synthesis of effects of plant species diversity on trophic groups and interactions. Nature Plants 6 (5), 503-510, DOI: 10.1038/s41477-020-0654-y.
- [60] Dassou A.G., Tixier P. (2016). Response of pest control by generalist predators to local-scale plant diversity. A meta-analysis. Ecology and Evolution 6 (4), 1143-1153, DOI: 10.1002/ece3.1917.
Butterfly and grasshopper communities were composed of common and generalist species with low species numbers, which contrasts with the high diversity of wild bees. However, these two taxa were surveyed in only one of the selected MAP fields. We assume that MAP fields are not equally suitable for each taxon. Differences in the suitability for different taxonomic groups of our three surveyed MAP fields were clearly visible, such as in the diversity of wild bees and Cerambycidae. While wild bees were the least diverse in one surveyed site (Prati/Wiesen), Cerambycidae were the most diverse at the same site. Climatic differences, on-field habitat and resource diversity and availability, and surrounding landscapes in particular may explain most of these patterns. According to several studies, landscape plays an important role in providing species with resources for reproduction and development [24][61]. In our study, the diversity of habitats at the landscape scale might be reflected in the different habitat requirements of bird and bat species detected in a sub-selection of MAP fields. Both taxa are highly mobile and therefore dependent on the surrounding landscape [24][35][62], which was mainly composed of grasslands, forests, urban settlements and at one site (Merano/Meran, M) intensive apple orchards, as well. The different habitats surrounding the MAP fields were well reflected by detected bird species pool. Several bird species, such as Phoenicurus ochruros (S.G. Gmelin, 1774) and Parus major (Linnaeus, 1758), breed in tree cavities and are therefore dependent on old trees found in forests and orchard meadows. In contrast, Turdus philomelos (C.L. Brehm, 1831), a bird species often found in intensive apple orchards, was only detected at the site where apple orchards were also present in the surroundings. Bird species adapted to urban environments, such as Pica pica (Linnaeus, 1758) and Passer italiae (Vieillot, 1817), were also found [63]. In terms of bats, ten species were detected at the site in M, of which six were also recorded while feeding and therefore actively choosing the MAP field as hunting habitat. In general, MAP fields are comparable to flowering and species-rich meadows, which provide a high number and diversity of insects throughout the growing season and are therefore good foraging habitats for bats [35]. The MAP field in M was located near a forest, which is why mainly edge-space foragers seemed to use the MAP field as a hunting ground. One bat species in particular, Rhinolophus hipposideros (Bechstein, 1800), stood out. This endangered species isn’t usually among those commonly observed, however, in our study it was the bat species with the most total calls. Moreover, it was the first time the species was recorded hunting during the four years of BMS surveys (Chiara Paniccia, unpubl. data).
×
- [24] Anderle M., Paniccia C., Brambilla M. et al. (2022). The contribution of landscape features, climate and topography in shaping taxonomical and functional diversity of avian communities in a heterogeneous Alpine region. Oecologia, 199, 499-512, DOI: 10.1007/s00442-022-05134-7.
- [61] Martin E.A., Dainese M., Clough Y., et al. (2019). The interplay of landscape composition and configuration. New pathways to manage functional biodiversity and agroecosystem services across Europe. Ecology Letters, 22 (7), 1083-1094, DOI: 10.1111/ele.13265.
- [35] Huemer P. (2004). Die Tagfalter Südtirols (Veröffentlichungen des Naturmuseums Suedtirol , 2). Folio Verlag, Vienna, Austria.
- [62] Tscharntke T., Grass I., Wanger T.C. et al. (2021). Beyond organic farming. Harnessing biodiversity-friendly landscapes. Trend in Ecology and Evolution 36 (10), 919-930, DOI: 10.1016/j.tree.2021.06.010.
- [63] Cramp S., Simmons A.D., Perrins C.M. (1994). Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palaearctic. Oxford University Press, Oxford, UK, here pp. 54, 288.
Overall, there is great potential for further research addressing biodiversity in MAP fields from multiple perspectives. Although the sampling effort was not equal for all MAP fields and taxa, our results show that the studied MAP fields functioned as a resource-rich oases for a variety of taxa, ranging from lower trophic levels such as herbivores and pollinators to birds and bats. We conclude that small-scale MAP cultivation may aid the conservation of various taxa and the promotion of biodiversity in a landscape dominated by agriculture.
Acknowledgements
This research was co-financed by the European Regional Development Fund through the Interreg Alpine Space Programme (LUIGI: Linking Urban and Inner-alpine Green Infrastructure, project number ASP 863). Furthermore, we thank all farmers for granting us access to the survey sites and Luciano Filippi (Cecina, Italy) for his great contribution in the identification of wild bees.
References
- [1] Máthé Á. (2015). Utilization/Significance of Medicinal and Aromatic Plants. In: Máthé Á (ed.). Medicinal and Aromatic Plants of the World. Scientific, Production, Commercial and Utilization Aspects. Springer, Heidelberg, Germany, here pp. 1-12, DOI: 10.1007/978-94-017-9810-5.
- [2] Ahad B., Shahri W., Rasool H. et al. (2021). Medicinal Plants and Herbal Drugs. An Overview. In: Aftab T., Hakeem K.R. (eds.). Medicinal and Aromatic Plants. Healthcare and Industrial Applications. Springer, Cham, Switzerland, here p. 2, DOI: 10.1007/978-3-030-58975-2_1.
- [3] Lubbe A., Verpoorte R. (2011). Cultivation of medicinal and aromatic plants for specialty industrial materials. Industrial Crops and Products 34 (1), 785-801, DOI: 10.1016/j.indcrop.2011.01.019.
- [4] Leaman D.J. (2008). Conservation and Sustainable Use of Wild-sourced Botanicals. Planta Medica, 74, 11, DOI: 10.1055/s-2008-1075152.
- [5] Shippmann U., Leaman D.J., Cunningham A.B. (2002). Impact of Cultivation and Gathering of Medicinal Plants on Biodiversity. Global Trends and Issues. In: Biodiversity and the Ecosystem Approach in Agriculture, Forestry and Fisheries. Satellite event on the occasion of the Ninth Regular Session of the Commission on Genetic Resources for Food and Agriculture, Rome, Italy, October 12-13, 2002. Food and Agriculture Organization of the United Nations, Rome, Italy.
- [6] Salomon I., Haban M., Otepka P. et al. (2018). Perspectives of small- and large- scale cultivation of medicinal, aromatic and spice plants in Slovakia. Medicinal Plants 10 (4), 261-267, DOI: 10.5958/0975-6892.2018.00041.2.
- [7] Allen D., Bilz M., Leaman D.J. et al. (2014). European Red List of Medicinal Plants. Publications Office of the European Union, Luxembourg, Luxembourg, here p. 4.
- [8] Südtiroler Bauernbund (ed.) (2014). Nischenkulturen als Erwerbsmöglichkeit. Chance und Herausforderung für die Südtiroler Landwirtschaft. Südtiroler Bauernbund, Bozen/Bolzano, Italy. Retrieved December 15, 2022, from https://issuu.com/effektgmbh/docs/sbb_broschu__re_nischenkulturen.
- [9] Šálek M., Hula V., Kipson M. et al. (2018). Bringing diversity back to agriculture. Smaller fields and non-crop elements enhance biodiversity in intensively managed arable farmlands. Ecological Indicators 90, 65-73, DOI: 10.1016/j.ecolind.2018.03.001.
- [10] Licata M., Maggio A.M., La Bella S. et al. (2022). Medicinal and aromatic plants in agricultural research, when considering criteria of multifunctionality and sustainability. Agriculture 12 (4), 529, DOI: 10.3390/agriculture12040529.
- [11] Hoppe B. (ed.) (2007). Handbuch des Arznei- und Gewürzpflanzenanbaus. Bd. 3: Krankheiten und Schädigungen an Arznei- und Gewürzpflanzen. SALUPLANTA e. V., Bernburg, Germany.
- [12] Pramsohler M., Gallmetzer A., Castellan A. et al. (2022). Erster Nachweis und molekularbiologische Bestimmung von Donus intermedius (Coleoptera: Curculionoidae) als Schädling bei Zitronenmelisse in Südtirol. Laimburg Journal 4, DOI: 10.23796/LJ/2022.003.
- [13] Nickel H., Blum H., Jung K. (2014). Verbreitung und Biologie der an mitteleuropäischen Arznei- und Gewürzpflanzen schädlichen Blattzikaden (Hemiptera: Cicadellidae, Typhlocybinae). Cicadina 14, 13-42, DOI: 10.25673/92235.
- [14] Meyer U., Blum H., Gärber U. et al. (2010). Praxisleitfaden Krankheiten und Schädlinge im Arznei- und Gewürzpflanzenanbau. Julius-Kühn-Institut Selbstverlag, Braunschweig, Germany.
- [15] Pobożniak M., Sobolewska A. (2011). Biodiversity of thrips species (Thysanoptera) on flowering herbs in Cracow, Poland. Journal of Plant Protection Research 51 (4), 393-398, DOI: 10.2478/v10045-011-0064-2.
- [16] Gupta S.K., Karmakar K. (2011). Diversity of mites (Acari) on medicinal and aromatic plants in India. Zoosymposia 6, 56-61, DOI: 10.11646/zoosymposia.6.1.10.
- [17] Venkatesh Y.N., Neethu T., Ashajyothi M. et al. (2022). Pollinator activity and their role on seed set of medicinal and aromatic Lamiaceae plants. Journal of Apicultural Research, Vol. ahead-of-print, DOI: 10.1080/00218839.2022.2080949.
- [18] Kumari B., Ravinder R. (2017). Pollination studies in Tagetes minuta, an important medicinal and aromatic plant. Medicinal Plants – International Journal of Phytomedicines and Related Industries 9 (2), 140-142, DOI: 10.5958/0975-6892.2017.00021.1.
- [19] Hilpold A., Anderle M., Guariento E. et al. (2023). Handbook - Biodiversity Monitoring South Tyrol. Eurac Research, Bolzano, Italy, DOI: 10.57749/2qm9-fq40.
- [20] Vrdoljak S.M., Samways M.J. (2010). Optimising coloured pan traps to survey flower visiting insects. Journal of Insect Conservation 16, 345-354, DOI: 10.1007/s10841-011-9420-9.
- [21] Droege S., Tepedino V.J., Lebuhn G. et al. (2010). Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conservation and Diversity 3 (1), 15-23, DOI: 10.1111/j.1752-4598.2009.00074.x.
- [22] Goulet H., Huber J.T. (eds.) (1993). Hymenoptera of the world. An identification guide to families. Canada Communication Group, Ottawa, Canada, here pp. 65-529.
- [23] Bibby C.J., Burgess N.D., Hillis D.M. et al. (2000). Bird census techniques. Academic Press, London, United Kingdom, here pp. 91-112.
- [24] Anderle M., Paniccia C., Brambilla M. et al. (2022). The contribution of landscape features, climate and topography in shaping taxonomical and functional diversity of avian communities in a heterogeneous Alpine region. Oecologia, 199, 499-512, DOI: 10.1007/s00442-022-05134-7.
- [25] Fornasari L., Mingozzi T. (1999). Monitoraggio dell'avifauna nidificante in Italia. Un progetto pluriennale sulle specie comuni. Avocetta - Journal of Ornithology, 23, 153-153.
- [26] Barataud M. (2015). Acoustic ecology of European bats. Species Identification and Studies of Their Habitats and Foraging Behaviour. Biotope & National Museum of Natural History, Paris, France, here pp. 104-263.
- [27] Teets K.D., Loeb S.C., Jachowski D.S. (2019). Detection probability of bats using active versus passive monitoring. Acta Chiropterologica 21 (1), 205-213, DOI: 10.3161/15081109ACC2019.21.1.017.
- [28] Hilpold A., Kirschner P., Dengler J. (2020). Proposal of a standardized EDGG surveying methodology for orthopteroid insects. Palaearctic Grasslands 46, 52-57, DOI: 10.21570/EDGG.PG.46.52-57.
- [29] Guariento E., Rüdisser J., Fiedler K. et al. (2022). From diverse to simple: butterfly communities erode from extensive grasslands to intensively used farmland and urban areas. Biodiversity and Conservation 32, 867-882, DOI: 10.1007/s10531-022-02498-3.
- [30] Hsieh T.C., Ma K.H., Chao A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods of Ecology and Evolution 7 (12), 1451-1456, DOI: 10.1111/2041-210X.12613.
- [31] Nieto A., Roberts S.P.M., Kemp J. et al. (2014). European Red List of bees. Publication Office of the European Union, Luxembourg, Luxembourg, DOI: 10.2779/77003.
- [32] Scheuchl E., Willner W. (2016). Taschenlexikon der Wildbienen. Alle Arten im Porträt. Quelle und Mayer, Wiebelsheim, Germany, here pp. 100-817.
- [33] Westrich P. (2018). Die Wildbienen Deutschlands. Ulmer, Stuttgart, Germany, here pp. 399-719.
- [34] Hellrigl K. (2003). Faunistik der Ameisen und Wildbienen Südtirols (Hymenoptera: Formicidae et Apoidea). Gredleriana 3, 143-208.
- [35] Huemer P. (2004). Die Tagfalter Südtirols (Veröffentlichungen des Naturmuseums Suedtirol , 2). Folio Verlag, Vienna, Austria.
- [36] Ceresa F., Kranebitter P. (2020). Lista Rossa 2020 degli uccelli nidificanti in Alto Adige. Gredleriana 20, 57-70.
- [37] Hilpold A., Wilhalm T., Kranebitter P. (2017). Rote Liste der gefährdeten Fang- und Heuschrecken Südtirols (Insecta: Orthoptera, Mantodea). Gredleriana 17, 61-86.
- [38] Rondinini C., Battistoni A., Teofili C. (2022). Lista Rossa IUCN dei Vertebrati Italiani 2022. Comitato Italiano IUCN e Ministero dell'Ambiente e della Tutela del Territorio e del Mare, Roma, Italia, here p. 31.
- [39] Dietz C., Kiefer A. (2016). Bats of Britain and Europe. Bloomsbury Publishing, London, United Kingdom, here pp. 104-105.
- [40] Origazzi A., Bardgett R., Barrios E. et al. (2016). Global Soil Biodiversity Atlas. Publication Office of the European Union, Luxembourg, Luxembourg, here pp. 28-65.
- [41] Bellmann H. (20063). Kosmos-Atlas Spinnentiere Europas. Kosmos, Stuttgart, Germany, here pp. 20-254.
- [42] Koch K. (1999). Carabidae - Micropeplidae. In: Die Käfer Mitteleuropas (Bd. E1). Spektrum Akademischer Verlag, Heidelberg, Germany.
- [43] Schöller M. (1996). Ökologie mitteleuropäischer Blattkäfer, Samenkäfer und Breitrüssler. Eigenverlag des EVCV, Bürs, Austria, here pp. 5-11.
- [44] Rheinheimer J., Hassler M. (2013). Rüsselkäfer Baden-Württembergs. Verlag Regionalkultur, Heidelberg, Germany, here pp. 31-67.
- [45] Klausnitzer B. (2011). Stresemann - Exkursionsfauna von Deutschland. Bd. 2: Wirbellose. Insekten. Spektrum Akademischer Verlag, Heidelberg, Germany, here pp. 111-316.
- [46] Harde K.W., Helb M., Elzner K. (2021). Der Kosmos Käferführer. Franckh-Kosmos Verlag, Stuttgart, Germany, here pp. 96-259.
- [47] Wilson J.D., Whittingham M.J., Bradbury R.D. (2005). The management of crop structure. A general approach to reversing the impacts of agricultural intensification on birds? Ibis 147 (3), 453-463, DOI: 10.1111/j.1474-919x.2005.00440.x.
- [48] O’Neill K.M., Fultz J.E., Ivie M.A. (2008). Distribution of adult Cerambycidae and Buprestidae (Coleoptera) in a subalpine forest under shelterwood management. The Coleopterists Bulletin, 62 (1), 27-36.
- [49] Sakalian V., Langourov M. (2004). Colour traps a method for distributional and ecological investigations of Buprestidae (Coleoptera). Acta Societatis Zoologicae Bohemicae 68, 53-59.
- [50] Klausnitzer B., Klausnitzer U., Wachmann E. et al. (2016). Die Bockkäfer Mitteleuropas. VerlagsKG Wolf, Magdeburg, Germany, here pp. 118-133.
- [51] Földesi R., Kovács-Hostyánszki A., Korösi A. et al. (2016). Relationships between wild bees, hoverflies and pollination success in apple orchards with different landscape contexts. Agricultural and Forest Entomology 18 (1), 68-75, DOI: 10.1111/afe.12135.
- [52] Roquer-Beni L., Alins G., Arnan X., et al. (2021). Management-dependent effects of pollinator functional diversity on apple pollination services: A response-effect trait approach. Journal of Applied Ecology 58 (12), 2843-2853, DOI: 10.1111/1365-2664.14022.
- [53] Ahrné K., Bengtsson J., Elmqvist T. (2009). Bumble Bees (Bombus spp) along a gradient of increasing urbanization. PLoS One 4:e5574, DOI: 10.1371/journal.pone.0005574.
- [54] Kuppler J., Neumüller U, Mayr A.V. et al. (2023). Favourite plants of wild bees. Agriculture, Ecosystems and Environment 342:108266, DOI: 10.1016/j.agee.2022.108266.
- [55] Schindler M., Diestelhorst O., Härtel S. et al. (2013). Monitoring agricultural ecosystems by using wild bees as environmental indicators. BioRisk 8, 53-71, DOI: 10.3897/biorisk.8.3600.
- [56] Höring T.F. (2014). How aphids find their host plants, and how they don’t. Annals of Applied Biology 165 (1), 3-26, DOI: 10.1111/aab.12142.
- [57] Singh R., Singh G. (2016). Aphids and their biocontrol. In: Omkar (ed.). Ecofriendly Pest Management for Food Security. Academic Press, Cambridge, Massachusetts, USA, here pp. 63-108.
- [58] Haddad N.M., Crutsinger G.M., Gross K. et al. (2010). Plant diversity and the stability of food webs. Ecology Letters 14 (1), 42-46, DOI: 10.1111/j.1461-0248.2010.01548.x.
- [59] Wan N.-F., Zheng X.-R., Fu L.-W. et al. (2020). Global synthesis of effects of plant species diversity on trophic groups and interactions. Nature Plants 6 (5), 503-510, DOI: 10.1038/s41477-020-0654-y.
- [60] Dassou A.G., Tixier P. (2016). Response of pest control by generalist predators to local-scale plant diversity. A meta-analysis. Ecology and Evolution 6 (4), 1143-1153, DOI: 10.1002/ece3.1917.
- [61] Martin E.A., Dainese M., Clough Y., et al. (2019). The interplay of landscape composition and configuration. New pathways to manage functional biodiversity and agroecosystem services across Europe. Ecology Letters, 22 (7), 1083-1094, DOI: 10.1111/ele.13265.
- [62] Tscharntke T., Grass I., Wanger T.C. et al. (2021). Beyond organic farming. Harnessing biodiversity-friendly landscapes. Trend in Ecology and Evolution 36 (10), 919-930, DOI: 10.1016/j.tree.2021.06.010.
- [63] Cramp S., Simmons A.D., Perrins C.M. (1994). Handbook of the Birds of Europe, the Middle East and North Africa. The Birds of the Western Palaearctic. Oxford University Press, Oxford, UK, here pp. 54, 288.
